Microbes to Mammals: Ecological Comparisons Across Scales and the Pitfalls of False Analogies
Because the life history of every species is at some level unique, the challenge in attempting to forge a comparative synthesis is finding instructive (as distinct from trite) commonalities. On one hand, if important differences are overlooked, the analogies are misleading and forced. Indeed, this must happen occasionally because what is truly ecologically important may become known only in hindsight. Francis Crick has said ... “There isn’t such a thing as a hard fact when you’re trying to discover something. It’s only afterwards that the facts become hard” (Judson 1979, p. 114). Boulding (1964) has cautioned about the dangers of false analogies in a world driven by the “rage for order”, where order or pattern is perceived but in reality does not exist. Importantly, however, he says further that (p. 36) ... “the remedy for false analogy is not no analogy but true analogy.”
A good case in point of seemingly empty coincidence is what became of Hutchinson’s (1959) famous observation that average individuals from sympatric, congeneric species showed regular differences in various morphological features such as length of proboscis in bumblebees or length of skull in squirrels (the so-called ‘Hutchinsonian ratio’ or ‘rule’). This was a potentially important contribution to the concept of character displacement, or the shift that occurs when two partly allopatric (nonoverlapping) species become sympatric (overlapping) in portions of their range.
The data he presented (see Hutchinson’s Table 1, p. 153) from various sources referred to skull dimensions in some mammals and the cul- men length (upper ridge of the bill) in birds. Hutchinson’s brief comment on this specific subject in the context of an extensive conceptual paper ended with the summary and conjecture that in general ... “where the species co-occur, the ratio of the larger to the small form varies from 1.1 to 1.4, the mean being 1.28 or roughly 1.3. This latter figure may tentatively [emphasis added] be used as an indication of the kind of difference necessary to permit two species to co-occur in different niches but at the same level of a food-web” (p. 152).
In an era when the role of competition as a force shaping communities was in its ascendancy, and despite Hutchinson’s caveats, these ratios quickly became elevated to the stature of a rule embedded in the literature, and supplemented by numerous subsequent reports from other authors. The ratios frequently were seized on ... “as prima facie evidence that communities were organized according to the principles of limiting similarity” (Eadie et al. 1987, p. 1). Horn and May (1977) showed later, however, that the rule also applied to such inanimate objects as an ensemble of musical instruments and the wheel sizes of children’s bicycles and tricycles.
The ratios are now generally regarded as being artifactual (Roth 1981), an inevitable outcome simply of a lognormal distribution in animal size with small variance in distribution (Eadie et al. 1987). The conditions apply to many animate and inanimate objects. While the ecological principles of character displacement and limiting similarity undoubtedly are valid, the underlying processes cannot meaningfully be captured in a size ratio. Likewise, the similarity between animal size and abundance patterns and those of car ‘species’ also raise a cautionary note with respect to conclusions that can be validly drawn (see discussion in Chap. 4 and Gaston et al. 1993).
On the other hand, sweeping generalities set at a level to avoid peculiarities of specific circumstances are usually so bland as to be trivial and uninformative. This dilemma is shared by model builders who compromise between accuracy and generality (see earlier quotation from Box and Draper 1987 footnoted in Chap. 6). As seen above, the problem is confounded by the intermingling of fact and dogma.
At what biological level does it make sense to compare the ecology of microorganisms and macroorganisms? As noted in Chap. 1, all organisms are quite similar in terms of their basic cellular function. Similarities among taxa diminish, as they do among other entities, as one moves through higher levels of systems organization (Table 8.1 and Slobodkin 1988; see also Zamer and Scheiner 2014).
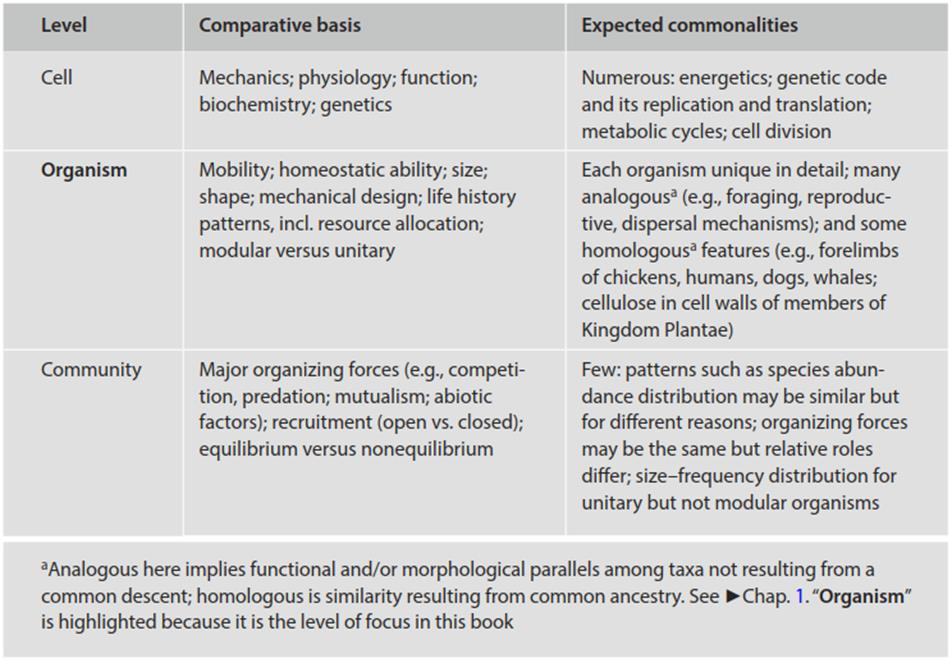
Table 8.1. Some levels at which all biota can be compared, the bases for comparison, and the anticipated similarities
This principle is but one of many examples of the general laws of integrative levels (Feibleman 1954): Each succeeding systems level embodies the attributes of its predecessors as well as imparting some of its own (emergent) properties. Hence, organisms can be expected to be more complex and distinctive as entities than the molecules of which they are mutually composed, and populations and communities, though sharing broad attributes, are yet more complex as systems than are the component organisms. For these and other reasons related to tractability and the levels on which natural selection effectively acts (see Preface and Introduction), the comparisons drawn in this book have been focused at the level of the individual. The complexities of ecological systems in many contexts, including the important implications of scale, are considered insightfully by Allen and Hoekstra (1992).
Date added: 2025-06-15; views: 49;