Urinary and Respiratory System. The Adult Salamander
The Adult Caecilian. The adult caecilian may have different excretory needs than the larva. In aquatic caecilians the role of the kidney is essentially unchanged (i.e., excretion of ammonia and water), whereas in the terrestrial caecilians the kidney may undertake the physiologically more demanding process of urea or uric acid excretion and water reab- sorption. (See Section 4.2, Water Homeostasis.)
The excretory system of the caecilian has been described (Wake, 1970a, b, 1972). The mesonephric kidney extends the length of the caecilian's coelomic cavity and retains the metameric arrangement of the larval pronephros (Plate 3.10). Caecilians lack the secondary tubules that arise in anurans and caudates. The mesonephros forms caudal to the pronephros but also functions by filtering both the coelomic and vascular fluids. The excretory route is similar to that described for the larval pronephros, however there are several changes. The nephrostome still connects with the coelomic cavity, but empties into a convoluted tubule. Vascular filtration is achieved by a Malpighian body, consisting of an internal glomerulus and Bowman's capsule, and this structure also empties into the convoluted tubule, which in turn empties into the mesonephric (common) duct. Many species of caecilians are known to have bladders that are bilobate. As with all amphibians, the mesonephric kidney cannot concentrate urine above the solute concentration of the plasma.
The dual filtration of coelomic and vascular fluid by the amphibian's mesonephric kidney may affect the distribution and clearance of intracoelomically administered drugs. Without specific studies documenting the pharmacokinetics of a given drug on a particular species, the clinician must rely on personal clinical judgment when administering intracoelomic drugs.
The Adult Salamander. The mesonephric kidney of the adult salamander arises posterior to the pronephros. The excretory route described for caecilians is similar to one family of caudates, the Amphiumidae, but secondary tubules are found in all other caudates. The secondary tubules arise at branch points of the primary tubules and may have glomeruli. Many tubules lack a nephrostome and are not connected to the coelomic cavity. Thus in many salamanders there is an emphasis on vascular filtration for excretion of nitrogenous wastes. However, as mentioned with caecilians, the role of the kidney is dependent on the Salamander's mode of life-most terrestrial salamanders will conserve water through the excretion of urea and reabsorption of water. The bladder is bilobate, bicornate or cylindrical in salamanders.
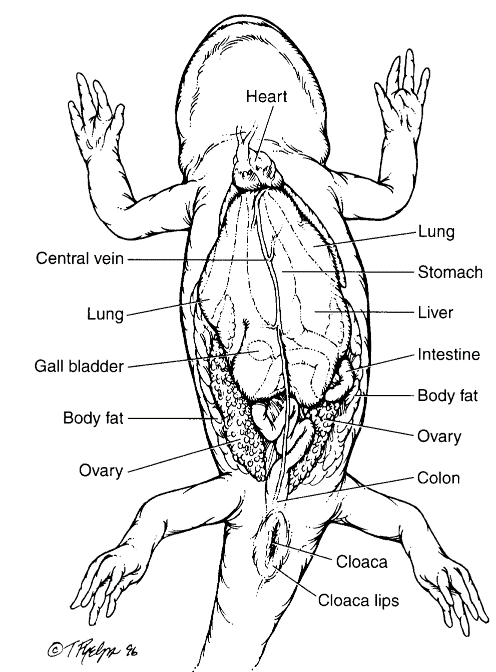
Figure 3.4. Visceral anatomy of a salamander. (Tim Phelps)
The Adult Anuran. The primary nitrogen waste of an anuran is either ammonia, urea, or uric acid, depending on its environment. Aquatic anurans excrete the majority of their nitrogenous waste as ammonia, since water conservation is not an issue. Many terrestrial species, such as bufonids, are not in constant con- tact with a water-rich environment and excrete a large portion of their nitrogenous waste as urea. Some anurans (e.g., African gray treefrog, Chiromantis xerampeiina, waxy treefrog, Phyllomedusa sauvagii) exploit relatively dry conditions by converting nitrogen wastes to uric acid. (See Section 4.2, Water Homeostasis.) Many of these uricotelic species possess cilia in the urinary bladder (Bolton & Beuchat, 1991).
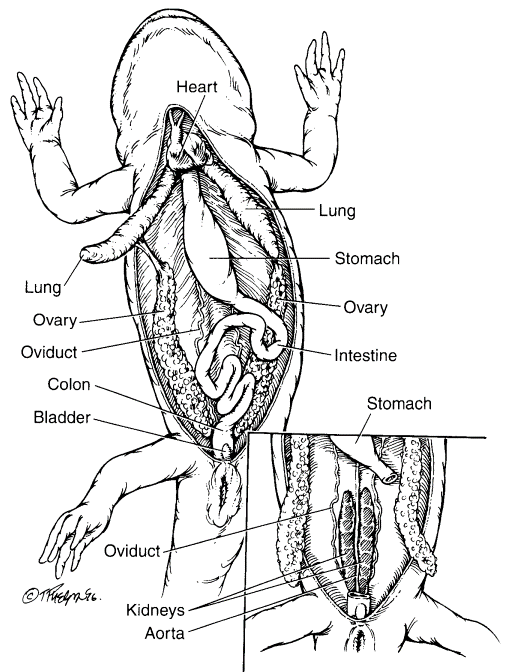
Figure 3.5. Visceral anatomy of a female salamander. Liver and gall bladder removed and right lung reflected. Inset view of retroperitoneal space. (Tim Phelps)
Respiratory System. The Adult Caecilian. There are three modes of adult respiration within the order Gymnophiona-pulmonic, buccopharyngeal, and cutaneous. The importance of each method undoubtedly varies based on the activity of the caecilian and the dissolved oxygen content of the water in which it lives. Little is known about the respiratory physiology of caecilians, and generalizations are impossible given the disparities in oxygen consumption under differing environmental conditions that have been documented for various species of salamanders and frogs.
The lung of the caecilian is the most important respiratory structure (Bennet & Wake, 1974). Most caecilians have paired elongate lung lobes, with the left lobe reduced in size or absent. In some species (e.g., typhlonectid caecilians), the right and left lobe are of approximately equal size. The alveoli of the caecilian lung is comprised of a mix of smooth muscle, cartilage and other connective tissue, blood vessels, and respiratory epithelium. In the aquatic typhlonectids, it is hypothe- sized that the lungs serve an additional role as an organ to maintain buoyancy as is noted for some aquatic salamanders (e.g., amphiumas, Amphiuma spp.).
Based on a study of a terrestrial caeciliid, the buccopharyngeal structures are the primary mechanical source for the inspiratory and expiratory effort (Mendes, 1945). The diameter of the external nares and the choana of a caecilian are controlled by smooth muscle. This musculature thereby limits the passage of air through the nasal duct into the buccopharyngeal cavity, and to some extent mediates olfaction by controlling access to the olfactory system.
The glottis is at the base of the tongue on the ventral aspect of the buccopharyngeal cavity, and in aquatic amphibians the glottis may be much smaller than expected for a similar sized terrestrial amphibian's glottis. The larynx leads immediately to the trachea. The trachea is well supported through its length with cartilaginous rings. The elongated trachea is lined with ciliated epithelium, and bifurcates into pulmonary bronchi. A tracheal lung has been described in some ichthyophiids and typhlonectids, and this structure actually serves as a structure for gaseous exchange, complete with respiratory epithelium.
The Adult Salamander. There are four modes of adult respiration within the order Urodela: branchial, cutaneous, buccopharyngeal, and pulmonic. The relative importance of any of these is dependent on the species in question. Neotenic species (e.g., sirens, Siren sp., mudpuppies, Necturus spp., axolotls, Ambystoma mexicanum, olm, Proteus anguinis, Texas blind salamanders, Typhlomolge rathbuni, etc.), as well as the aquatic larvae of many species, possess external gills and rely heavily on branchial respiration. Some aquatic species (e.g., sirens, Siren spp.) possess lungs in addition to their gills.
The structure of the gill is dependent on the environment of the species, and the shape and resultant surface area is, to a degree, mutable within a species, dependent on physical parameters. Aquatic environments with a high dissolved oxygen content allow salamanders to have short gills, whereas in areas of low dissolved oxygen content the gills tend to become long and elaborate. Stream-dwelling species tend to have small gills to reduce the drag of the current. The concomitantly high dissolved oxygen content of this environment permits this necessarily small surface area of the gill structure.
Cutaneous respiration is present to a variable degree in all species. This mode of respiration is feasible in salamanders as a result of several anatomic factors: high surface area to volume ratio resultant from the small body size and cylindrical shape, a thin epidermis, and a highly vascularized dermis. All of these features promote the exchange of gases across the skin. Coupled with a salamander's low metabolic rate and its ability to incur an oxygen debt through anaerobic glycolysis, these anatomic features permit cutaneous respiration. In fact, this mode of respiration is so successful that the plethodontids, one of the most speciesrich families of salamanders, are lungless.
The surface area to volume ratio is enhanced by the presence of lateral folds of skin in the aquatic cryptobranchid salamanders (e.g., hellbender, Cryptobranchus alleganiensis, Asian giant salamanders, Andrias spp.), and a rocking motion helps increase the diffusion rate across the skin by keeping a current of low dissolved carbon dioxide content water running across the skin. Costal grooves also serve to enhance the surface area of a salamander. The same characteristics that allow cutaneous respiration also play a role in the distribution of topically applied drugs.
The buccal cavity and pharynx serve as a site of gaseous exchange in many species of salamanders. This gaseous exchange is to some extent an extension of the cutaneous respiration, but the muscular pumping action to ventilate this region, as well as peculiarities of the vascular supply of these regions, support buccopharyngeal respiration as a distinct entity. The pumping action is the main source of both buccopharyngeal and pulmonic ventilation for the salamander, both inspiratory and expiratory.
Some salamanders are a ble to make noises using different anatomic features. The Pacific giant salamander, Dicamptodon ensatus, has vocal folds (Maslin, 1950), but the voice of salamanders is derived by forcing air through gill slits or nares or forming a vacuum during inspiration and then opening the mouth.
The lungs of salamanders vary in size and degree of partitioning among species. As a rule, the right and left lung are of approximately equal size, although the right side is slightly smaller than the left. The lungs are generally simple structures with no partitioning or in- folding in some aquatic salamanders (e.g., mudpuppies and waterdogs, Necturus spp.), while terrestrial salamanders tend to have sacculations and even alveoli in the anterior portion of each lung. Many pond- dwelling species have alveoli too. Although plethodontid salamanders lack lungs, in other families such as the hynobiid clawed salamanders, Onychodactylus spp., lungs are reduced in size, or even absent.
The trachea leads to a short bifurcation into the lungs. Cartilaginous rings are present in the trachea, and in some species, the bronchi. The trachea is relatively short in most salamanders, a fact which must be borne in mind when intubating for tracheal washes or intratracheal administration of drugs, and during gaseous anesthesia, or else the intubating device could be inserted too far and damage the pulmonic epithelium. Surprisingly the trachea of many aquatic salamanders, notably the amphiumas, Amphiuma spp., is greatly elongated as a result of the position of the lungs, due to the dual functions of gaseous exchange and hydrostasis.
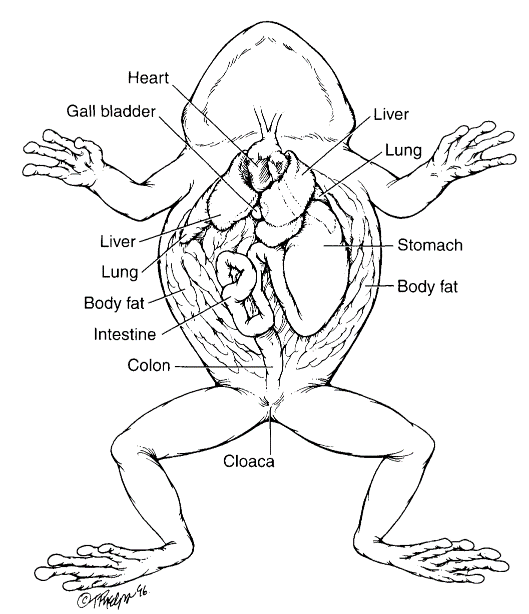
Figure 3.6. Visceral anatomy of a frog. (Tim Phelps)
The Adult Anuran. There are three modes of adult respiration within the order Anuracutaneous, buccopharyngeal, and pulmonic. The relative importance of any of these is dependent on the species in question.
The aquatic Titicaca water frog, Telmatobius culeus, has prominent folds of skin to increase its surface area, and relies heavily on cutaneous respiration, whereas the African clawed frog, Xenopus laevis, lacks folds and is primarily a lung breather. This is probably reflective of the dissolved oxygen content of the water in which these species evolved, with the cold well-oxygenated water of the mountain lake allowing the Titicaca water frog, Telmatobius culeus, to obtain adequate oxygen saturation of the blood through cutaneous respiration. This option is not available in the warmer waters in which the clawed frog, Xenopus laevis, is found.
The buccal cavity and pharynx serve as a site of gaseous exchange in many species of anurans. As in salamanders, buccopharyngeal respiration is a distinct mode of respiration in anurans. The pumping action is the main source of both buccopharyngeal and pulmonic ventilation for the anuran, both inspiratory and expiratory.
Anurans are the most vocal amphibians, with almost every species having a voice. The voice of anurans is derived by forcing air from the lungs across the vocal slits and into the vocal sacs by a combination of buccopharyngeal pumping and pulmonic pumping.
The trachea is extremely short and bifurcates in the lungs. Cartilaginous rings are present in the trachea. The shortness of the trachea must be considered when the clinician is intubating for tracheal washes or intratracheal administration of drugs, and during gaseous anesthesia, or else the intubating device could be inserted too far and damage the pulmonic epithelium.
The lungs of anurans vary in size and degree of partitioning among species. As a rule, the right and left lung are of approximately equal size. The lungs are generally simple structures with no partitioning or in folding.
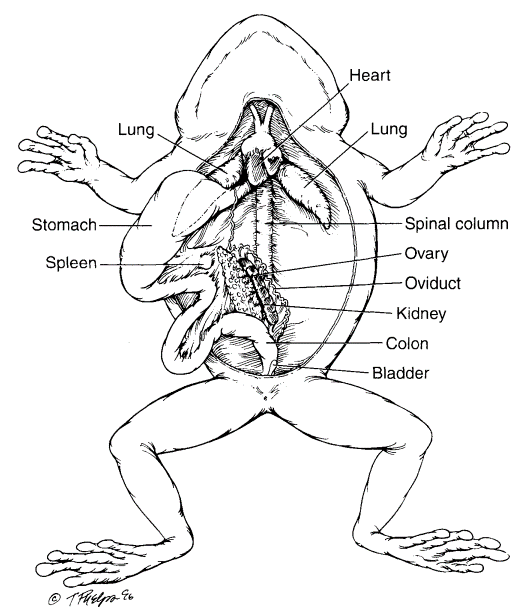
Figure 3.7. Visceral anatomy of a female frog. Liver and gall bladder removed and stomach reflected to reveal urogenital system. (Tim Phelps)
Date added: 2022-12-11; views: 1291;