Coordination of Movement Direction, Muscle Activation in Isometric Contractions
One of the main problems concerning the coordination of muscle activation for movements or torques in a particular direction is that for some motor tasks, the number of muscles acting across a particular joint is larger than strictly necessary. For example, there are at least seven muscles acting across the elbow joint, contributing to flexion/extension and supination/pronation (i.e., rotation along the longitudinal axis of the forearm).
This gives rise to the possibility that a torque or movement in a particular direction can be realized by a large variety of different muscle activation patterns. Despite this redundancy, a unique activation pattern of muscles is observed in various subjects for isometric contraction in a particular direction. Note that we do not imply that the motor system is redundant in general, because different muscles may have muscle fibers with different histochemical properties and with a different fiber architecture, which gives each muscle particular functional properties.
A. Muscle Activation in Isometric Contractions. To investigate the activation of muscles in detail, the activation of single motor units in human arm muscles has been measured. Muscle force is graded by two mechanisms: recruitment (i.e., the increase of the number of active motor units) and firing rate of motor units. The relative contribution of each of these mechanisms is different for proximal (i.e., closer to the center of the body) and distal (i.e., farther away from the center of the body) muscles, with a more prominent role for recruitment in proximal muscles and a more prominent role for rate modulation in distal muscles.
The motor units in a muscle all have a different recruitment threshold in the range from very small to very large forces. Because a motor unit is thought to be recruited every time the input to that motoneuron exceeds a particular level, the recruitment threshold of a motor unit may be considered as a condition in which the total synaptic input to that motor unit is constant. This phenomenon was used to study the input to motoneurons in different experimental conditions.
The human biceps brachii is frequently called a multifunctional muscle because m. biceps contributes to torques in flexion and supination direction. This multifunctional character is reflected in the activation of motor units. Figure 1 shows the recruitment threshold of several motor units in m. biceps brachii, caput longum. For the motor units in Fig. 1A, the recruitment threshold depends on the torque in flexion direction only. Whatever the torque in supination direction, the recruitment threshold for flexion remains the same. This indicates that motor units of this type are activated for flexion torques in the elbow.
Figure 1B shows the recruitment behavior of another type of motor units in m. biceps, caput longum. The recruitment threshold of motor units of this type depends on the torque in supination direction, whatever the torque in flexion direction.
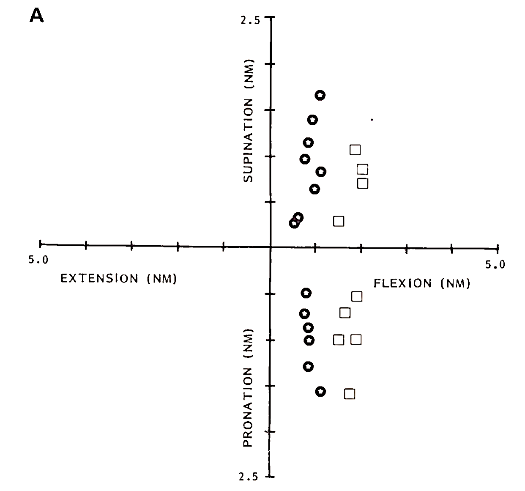
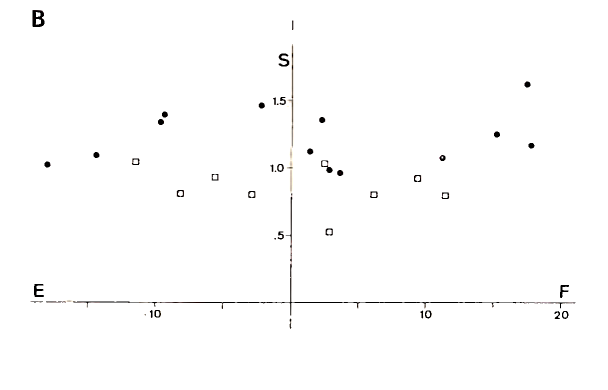
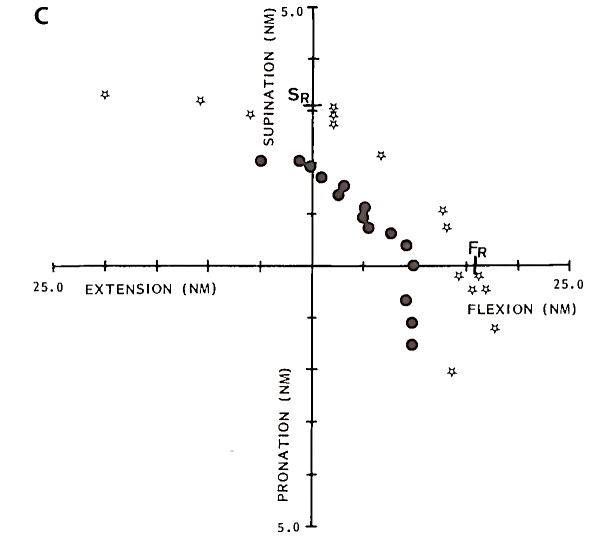
FIGURE I. Example of motor-unit behavior in m. biceps for three motor-unit subpopulations. With different symbols (squares, circles, and stars), recruitment behavior is illustrated for two units. Each symbol indicates the combination of torques at which a motor unit is recruited. Recruitment threshold is plotted in Newton meters (Nm) in flexion (F), extension (E), supination (S), and pronation.
For one motor unit (C), the recruitment threshold for flexion and supination direction is indicated along the horizontal and vertical axis, respectively. (A, B) Recruitment behavior of motor units, the recruitment threshold of which depends only on flexion torque or supination torque, respectively. (C) Data are shown for two motor units for which the recruitment threshold depends on torques in both flexion and supination direction
The majority of motor units in m. biceps, caput longum, are recruited for torques in both flexion and supination direction. An example of this type of behavior is shown in Fig. 1C. All recruitment thresholds fall along a straight line in the flexion/supination plane, which can be described by the equation
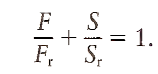
In this formula, F and S refer to the torque in flexion and supination direction, respectively, and Fr and Sr refer to the recruitment threshold in flexion and supination direction, respectively. For extension, the recruitment threshold of these motor units depends on the torque in supination direction only, and for pronation, the recruitment threshold depends on the torque in flexion direction only.
The combinations of torques in Fig. 1C, where the two motor units are recruited, fall along parallel lines. This is related to the fact that the ratio of recruitment thresholds in human biceps muscle is the same for all motor units of this type. This is illustrated in Fig. 2, where Fr and Sr are plotted for a representative sample of motor units in m. biceps brachii. The fact that all the data points in Fig. 2 fall approximately along a straight line indicates that the input to all motor units of this type is the same. Motor units may have a different recruitment threshold, but the relative input related to flexion and supination is the same for all motor units of the same type. The results in Fig. 1 are very consistent and reproducible over various subjects and indicate that the coordination of muscles for torque in a particular direction is unique except for a cocontraction term of antagonistic muscles.
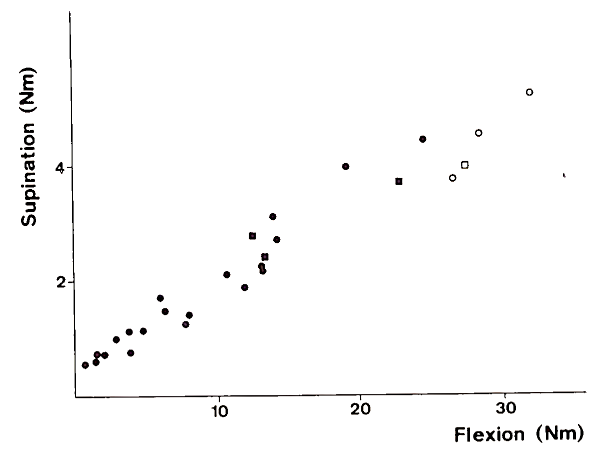
FIGURE 2. Recruitment threshold [in Newton meters (Nm)] for flexion and supination for motor units of the type shown in Fig. 1C. Circles and squares refer to data of motor units in the long head and short head, respectively, of m. biceps. The fact that all data points tend to fall along a straight line suggests that the ratio of recruitment thresholds in flexion and supination direction is constant
The motor units of a particular subtype are not randomly distributed in the muscle but are clustered in a specific location of muscle. In fact, there is a specific relation between location of a motor unit in muscle and its recruitment behavior. This is illustrated in Fig. 3, which shows the intramuscular EMG activity recorded with intramuscular wire electrodes in the lateral and medial side of m. biceps brachii. When flexion torque is increased, activity at the lateral side increases, and when flexion torque decreases and supination torque increases, the center of activity shifts from the lateral to the medial side of m. biceps brachii.
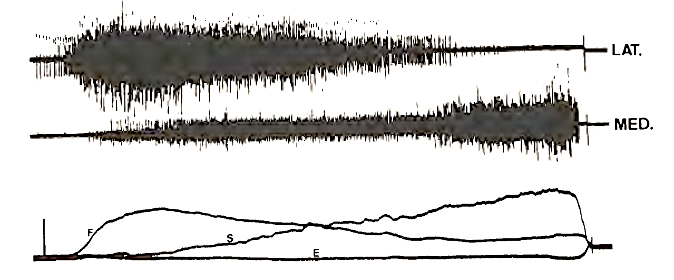
FIGURE 3. Intramuscular recordings of electromyographic activity obtained at the lateral side (top trace) and medial side (middle trace) in m. biceps, caput longum, when isometric flexion torque is initially increased and gradually decreased while supination torque is gradually increased. Lower trace shows torques in flexion (F), supination (S), and exorotation (E). Exorotation is a rotation in the shoulder joint along a longitudinal axis along the upper arm
These results indicate that the activation of the motoneuron population of m. biceps brachii is inhomogeneous. However, for each subpopulation the results are compatible with the notion of a homogeneous activation of a particular pool of motoneurons. The idea of homogeneous activation is related to the notion of the size principle. According to this principle, the same input is received by each of a set of motoneurons, and the recruitment threshold of each motoneuron is related to the size of the motoneurons cell body.
This idea predicts that the recruitment order of motoneurons is the same in all conditions. Without further arguments, the size principle has been generalized in the literature to the whole motoneuron pool of a muscle, although this is not implied by the strict definition of the size principle. The data in Fig. 1 clearly reveal an inhomogeneous activation of the motoneuron pool of m. biceps, and similar data have been presented for almost all muscles. However, these data do not argue against the size principle.
Rather the data in Fig. 1 suggest that the motoneuron pool of a muscle has several groups and that each group receives a distinct but homogeneous activation. Recent experiments in animals have confirmed the compartmentalization of the motor-unit pool in a single muscle in several subpopulations and have shown that the motoneurons of motor units of the same subtype are clustered together in the spinal cord. This observation suggests that for movements or torques in different directions, different locations in the spinal cord may be activated.
The compartmentalization of motor units of a particular type in a specific part of muscle has important methodological implications because numerous studies have used surface EMG activity as a measure of muscle activation. Because EMG activity recorded with surface electrodes reflects the weighted activity of some part of the muscle fibers, the EMG activity strongly depends on the position of the surface electrodes relative to the muscle. Therefore, EMG activity recorded with surface electrodes cannot provide a complete picture of muscle activation.
Because motor units that were active during flexion and supination were localized in different parts of m. biceps, the question arose whether motor units in different subpopulations had a different mechanical advantage. The mechanical advantage of a muscle is the mechanical effect of muscle force to the force exerted by the limb. Evidently, this mechanical advantage is optimal when the pulling direction of muscle is orthogonal to the bone of insertion. To investigate this issue, the contribution of motor-unit twitches in supination and flexion direction was determined. Because motor units may have a different twitch amplitude, the argument was that if all motor units contribute equally to flexion and supination torques, the ratio of twitches in flexion and supination direction should be the same for all motor units.
Any systematic violation of a constant ratio for motor units in different subpopulations would suggest a relation between recruitment behavior and mechanical advantage. It appeared that no relation whatsoever exists between the ratio of twitches in flexion and supination directions for motor units in human arm muscles. This indicates that the compartmentalization in m. biceps reflects a neural organization rather than a difference in mechanical advantage.
An inhomogeneous activation of the population motor units has been found in almost all muscles involved with flexion/extension and supination/protonation. It has also been found in muscles that were not considered multifunctional, such as m. triceps, which generates a torque in extension only. For a small sample of motor units in m. triceps, the recruitment threshold depends on torque in extension direction only. However, there is a larger group of motor units in m. triceps that is also activated for torques in supination and pronation direction.
The activation for torques in supination direction can be understood from the fact that m. biceps is activated for torques in supination direction. Because m. biceps has a mechanical advantage with a component in flexion direction, a torque in supination direction can be obtained only if m. triceps compensates for the torque component in flexion direction by m. biceps. The activation of motor units in m. triceps for pronation can be understood from the contribution of m. pronator teres to torque in pronation and flexion direction.
The mechanical advantage of muscle changes as a function of joint angle. This is illustrated in Fig. 4, which shows the combinations of torque where a single motor unit in m. biceps is recruited at different flexion angles in the elbow joint. Clearly the ratio of recruitment thresholds in flexion and supination direction, FJSr, changes as a function of joint angle. Similar changes in recruitment behavior have been observed in nearly all human arm muscles. These results indicate that the coordination of muscles and the precise distribution of muscle activations are functions of joint angle.
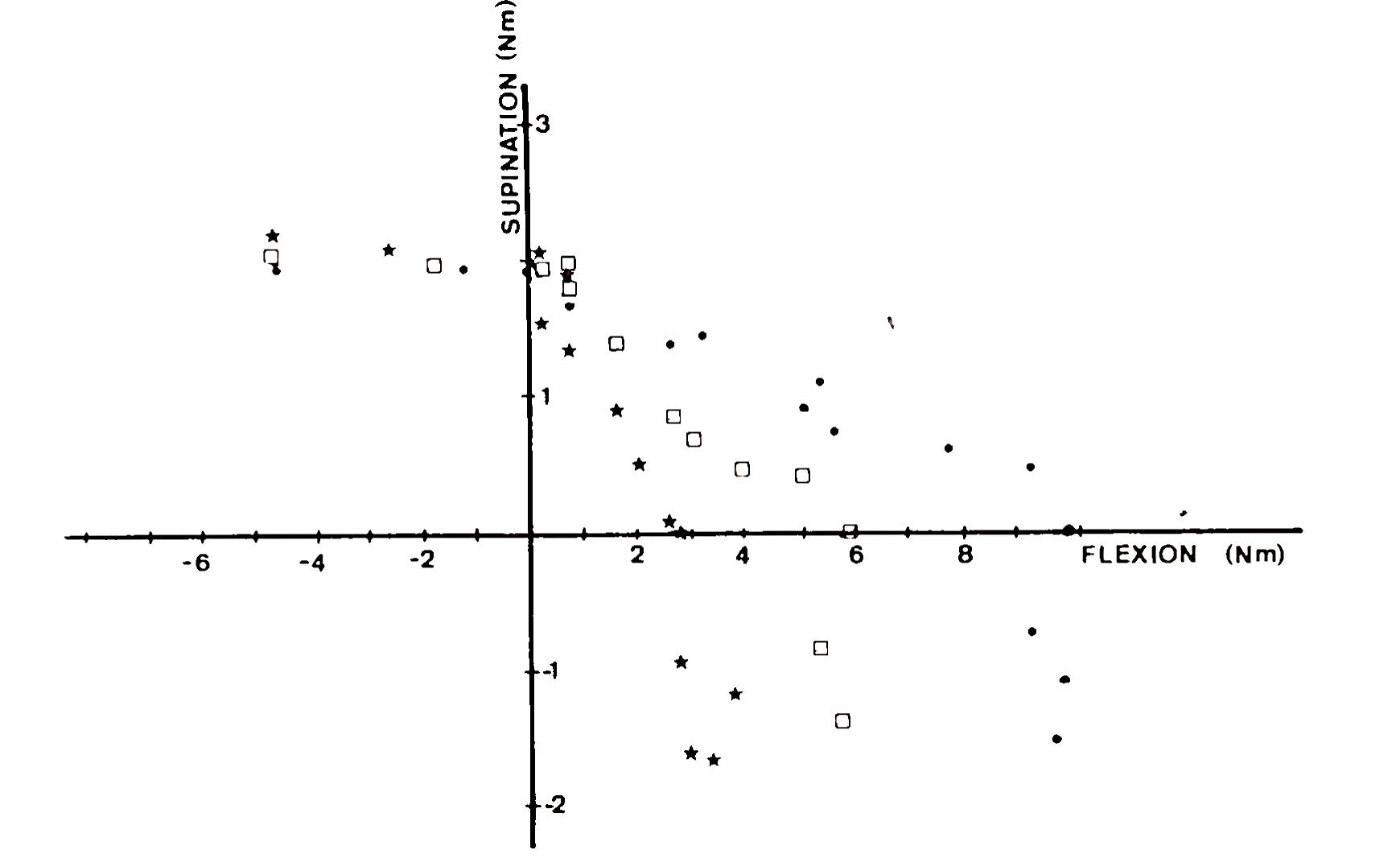
FIGURE 4. Recruitment threshold in Newton meters (Nm) for a single motor unit in m. biceps brachii (caput longum). Each data point indicates the recruitment threshold for a combination of forces in flexion/extension and supination/pronation. Different symbols refer to data obtained at different angles between forearm and upper arm: 100° flexion (circles), 145° flexion (squares), and 175° flexion (stars)
To provide an explanation for the inhomogeneous activation of the motor-unit pool of muscle, the concept of task group has been proposed, in which functional groups of alpha and gamma motoneurons and spindle afferents are programmed to achieve optimal control of movements. Alpha motoneurons are the motoneurons that activate muscle fibers and thereby produce force. The gamma motoneurons activate the muscle spindles, thereby regulating their sensitivity. The problem with the notion of task group is that the criterion for “optimal” is not always properly defined. However, several suggestions have been made:
- Compartmentalization due to a different mechanical advantage. Some muscles have a broad attachment at the tendon such that different muscle fibers may have a different mechanical advantage (e.g., in cat sartorius muscle).
- Compartmentalization due to a different functional role. For example, some biarticular muscles are activated during both shortening and lengthening. In these two conditions, different motor-unit groups may be activated.
- Compartmentalization due to different mechanical properties of muscle fibers. For some muscles (e.g., gastrocnemius), muscle fibers in different muscle locations have different properties with respect to twitch contraction time, fatigability, and twitch amplitude.
- Compartmentalization reflecting a neural organization, such as in the human m. biceps brachii.
Presumably, all of these mechanisms may underlie the compartmentalization of muscle in general.
Date added: 2022-12-11; views: 886;