Weather Impact on Vultures Migration: Climate, Thermals, and Flight Strategies
Weather conditions also have a strong affect on the migration of birds. Local weather conditions affect, among others, the start and duration of migration, energy cost, migratory routes, flight speed and flight strategies of raptors (Maransky 1997; Shamoun-Baranes et al. 2006; Shamoun-Baranes 2010; Vardanis 2011). Apart from temperature, other factors for migration are availability of food, day length and breeding. Studies have found that migrating birds use the sun and stars, the earth's magnetic field, and mental maps for direction. In some species, not all populations are migratory; this is termed partial migration.
Shamoun-Baranese et al. (2010: 280) point out that 'meteorology should be integrated into research on migration, especially when trying to understand natural variability observed in aspects like the timing of migration; migratory routes; orientation; use of stopover sites; or population trends such as effects on survival or breeding success as a result of changes in arrival time or physiological condition.' They further note the importance of the longer term meteorological dynamics, namely climate; 'meteorology is also of interest at longer time scales when trying to understand the evolution of particular migratory systems' (ibid.).
These authors list some recent, important literature that provides a background for further discussion of this topic for general lists of bird species. For general overviews, see Richardson (1978, 1990), Drake and Farrow (1988), Dingle (1996), Liechti (2006), Newton (2008). Other topics concern the impact of atmospheric conditions on migration onset (Shamoun- Baranes et al. 2006; Gill et al. 2009), migratory success (Erni et al. 2005; Reilly and Reilly 2009), flight speeds (Garland and Davis 2002; Shamoun-Baranes et al. 2003), flight altitudes (Bruderer et al. 1995; Schmaljohann et al. 2009), flight strategy (Pennycuick et al. 1979; Spaar and Bruderer 1997; Spaar et al. 1998; Sapir 2009), bird stopover decisions (Akesson and Hedenstrom 2000; Danhardt and Lindstrom 2001; Schaub et al. 2004), migration phenology (Huppop and Huppop 2003; Jonzen et al. 2006; Bauer et al. 2008, 2009), flight orientation and flight trajectories (Thorup et al. 2003), and the probability and/or intensity of migration (Erni et al. 2005; van Belle et al. 2007).
Medium and large birds, with wing loadings that are relatively high, tend to migrate by soaring, using horizontal or vertical (thermal) winds rather than active flight (Alerstam 1990). Bohrer et al. (2012: 96) investigated the migratory behavior and the meteorological conditions for thermal and orographic uplift 'to contrast flight strategies of two morphologically similar but behaviorally different species: the Golden eagle, Aquila chrysaetos, and Turkey vulture, Cathartes aura, during autumn migration across eastern North America.' They argued that 'it has not been shown previously that migration tracks are affected by species-specific specialization to a particular uplift mode' (Bohrer et al. 2012).
This study noted that both the Golden eagle and the Turkey vulture are soaring migrants that use both thermal and orographic uplift. Differences involved the higher wing loading (mass/wing area) of golden eagles (7.2 kg m2) than the turkey vultures (3.9 kg m2) (see also Pennycuick 2008). Golden eagles have 'more muscle mass and thus stronger flapping flight than turkey vultures.' The Turkey vulture's lower wing-loading is adapted to the use of weak updrafts and a scavenging (non-predatory) specialization, while the higher wing loading of golden eagles is for faster flight and a predatory specialization. The Golden eagle's preference for strong orographic lift favored flight paths above the windward side of the Appalachian mountains and ridges (see also Allen et al. 1996; Brodeur et al. 1996). By contrast, Turkey vultures used thermals, even when the sources of the thermal lift were scattered (Boehrer et al. 2011, see also Mandel and Bildstein 2007).
For the use of thermals for migration, problems emerge when they have to cross the ocean, as thermals form only over land. This results in large numbers of birds using 'bottlenecks'; narrow land links between continents (e.g., the isthmus of Panama) and narrow seas that partition landmasses (e.g., Gibraltar, Falsterbo, and the Bosphorus) (Fig. 6.3). Other migratory links include passes between high mountains; examples are the Central American migratory bottleneck and the Batumi bottleneck in the Caucasus, which allows avoidance of the vast Black Sea and high mountains (Maanen et al. 2001).
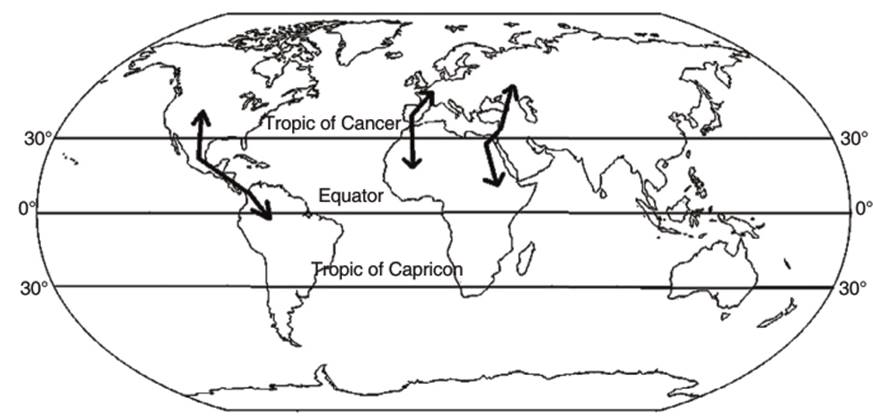
Fig. 6.3. Migration Bottlenecks for Raptors
For the New World vultures, a crucial flyby is the Mesoamerican Land Corridor, connecting North America and South America, 'numerically the most important raptor-migration flyway in the world' (Bildstein and Sabario 2000: 197). These authors report that Buteo platypterus made up 76%, B. swainsoni 14%, and Cathartes aura 10% of one sample (see also Carriker 1910; Skutch 1945; Andrle 1968; Smith 1980, 1985a, 1985b; Thiollay 1980; Stiles and Skutch 1989, Tilly et al. 1990; Hernandez and Zook 1993; Hidalgo et al. 1995; Sutton and Sutton 1999; Zalles and Bildstein 2000). In this corridor, most birds 'soared in and glided between thermals; some engaged in slope soaring' (Bildstein and Sabario 2000: 200).
The results of this study suggested that migration is more likely on still, raining and overcast times in Costa Rica than in temperate regions. One study of migratory Swainson's hawks migrating between Argentina and North America show that they traveled 42% faster in the tropical part of the journey than in the temperate portions (Fuller et al. 1998); hence the dynamics of raptor flight may differ between tropical and temperate regions (Bildstein and Sabario 2000).
Griffon vultures have been documented using migratory bottlenecks, such as the Borsporus and Suez for migration (Bernis 1983). Bildstein et al. (2009: 382) studying the migrating flight behavior of Griffon vultures (Gyps fulvus) at another major migration bottleneck, the Strait of Gibraltar, found that the 14-km-wide sea channel 'significantly impeded the southern migration of the species into Africa, with many birds attempting repeated passage for weeks before crossing, and others not crossing at all and overwintering in Southern Spain.' Most attempts to cross the straits were between 11:00 and 14:00 hours, when there were either light or variable winds, or strong winds from the north or west. The study found that there were no crossing attempts when strong winds blew from the east or south. Furthermore, Griffons never tried to cross the straits alone; attempts were only made in large flocks.
Flight modes over water varied from those over land; above water, birds flapped more than ten times more often, and also did more flaps per flapping than when flying over land before the crossing attempts. The flapping was intermittent, combined with gliding or soaring flight. An important finding was that 'vultures flying over water that flapped at rates of 20 flaps or more per minute typically aborted attempted crossings and returned to Spain in intermittent flapping and gliding flight' (ibid. 382).Exhausted vultures fell into the sea and drowned and also collapsed on the beach after the crossing (see also Barrios Partida 2006). This indicated that Griffon vultures could not maintain flapping flight for even a short time, and that even narrow straits are thus serious barriers to migrating vultures.
Another study in another popular population bottleneck in Israel (Leshem and Yom-Tov 1998) found that migrating soarers use three main topographically determined routes in both directions in the autumn and the spring; an eastern route along the western edge of the central mountain range; an eastern route along the Jordan Valley and the Dead Sea towards the Sinai, merging with the western route in autumn; and a southern-Elat mountains route. Wind was also important, as in the morning breeze from the Mediterranean Sea shifted the migration path eastwards. This was also found to deflect the migration of the eastern route in the autumn. This route is favored when it moves over the eastern side of the central mountain range during the morning, as the sun creates thermals for soaring; in the afternoon the sea breezes deflect the path eastwards to the Jordan Valley. In the Elat mountains, the wind is affected by the southern Arava valley which moves the path in a north-south direction. Seasonally, it has been noticed that early autumn migrants move on an eastern route, but later migrants fly along the Mediterranean coast, possibly due to 'sub-optimal soaring conditions along the coastal plain during August' (ibid. 41). This is probably due to temperature differences between the land and sea, these differences declining and then disappearing by October. Species differences in routes were attributed to the avoidance of the Red Sea (Leshem and Yom-Tov 1998).
Another study, also focusing on this migration bottleneck in Israel (Spaar 1997) looked at the flight styles of migrating raptors. There was very little variation in the climbing rate of different species in thermal circling. The strength of thermal updrafts was the main determinant. Also, in interthermal gliding, heavier species had smaller gliding angles and glided faster, as gliding ability increased, than in lighter species. The Griffon vulture (Gyps fulvus) used soaring in a straight line whilst gliding. The Egyptian vulture was a soaring migrant.
Seasonal climate change may also contribute to mammal migration. This may influence movements of vultures that follow migratory mammals for food. For example, as noted above, Griffon vultures being pure scavengers follow migratory herds of ungulates in the African savannas, unlike the smaller, more adaptive Hooded and Egyptian vulture, which must find smaller prey items when the ungulates migrate. In the past colonial times, migratory herds were much larger than at present (Meinertzhagen 1957; Dorst 1970). In India as well, there were large migrations of the Blackbuck Antilope cervicapra during the 19th century (Blanford 1907; Seshadri 1969). These migrations may have shaped the migratory foraging behavior of the Griffons. Vultures are dependent on ungulate mortality, due to disease, accidents, old age and starvation and these are highest towards the end of the dry season. This is due to the lowered protein content of the foraged vegetation, which contributes to a decline in the strength and health of ungulates (see Sinclair 1977; Houston 1983).
The ungulate herds follow the movement of the Intertropical Convergence Zone. This zone is based on the movements of the sun and rain north across the Equator in April, May and June, staying north of the equator from July to September, and then moving south of the Equator again from October to December. This affects vultures, as ungulates are stronger during the rains, so vultures must follow the location of the period towards the end of the dry season, which varies in location. For example, the dry season in the Serengeti continues until August-October, when the rains slowly arrive as the Intertropical Convergence Zone moves south and the November rains begin. At this time the dry season is in Sudan. There, if vultures could fly distances, they could exploit weaknesses of the ungulates brought on by the regional occurrence of the dry season. This would mean a north-south migration, just in front of the rainbelt (Houston 1983). This study notes that in southern Africa, the seasonal rainbelt moves in a northwest-southeast line. The Griffons would have optimal feeding in the southeast (e.g., Zululand) in the dry season of June to September, and in southern Angola during the dry season of October to December.
Date added: 2025-04-29; views: 376;