Amphibian and Reptile Natural History
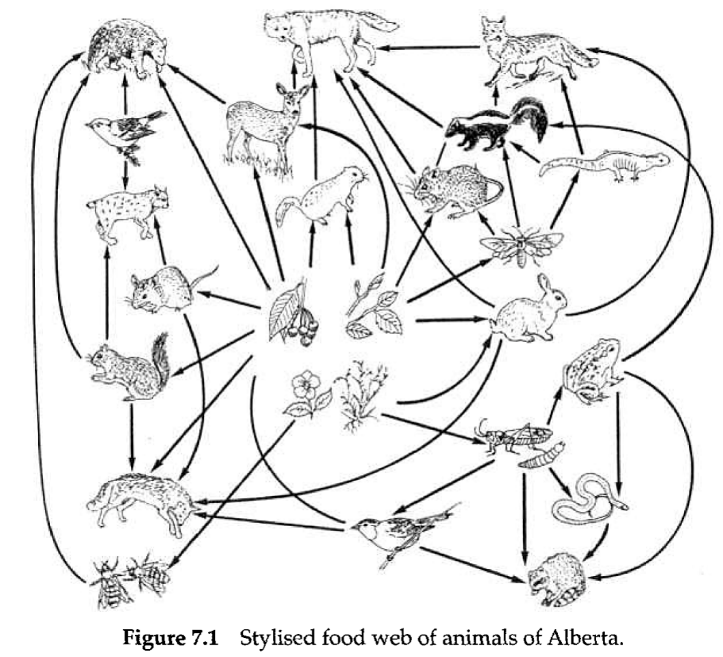
REFERENCE TO THE SPECIES accounts in Chapter 5 clearly indicates that amphibians and reptiles interact with a wide variety of vertebrates and invertebrates in relation to trophic dynamics. The herpetofauna thus plays a key role in the food webs of a variety of communities (Fig. 7.1). Indeed, amphibian and reptile consumption of insects and rodents, respectively, may be far greater than generally appreciated. Likewise, amphibians, especially, contribute a significant resource base for both terrestrial and aquatic predators. A single species of amphibian will participate in at least two community trophic relation systems.
Tadpoles are generally herbivores and occur in huge numbers, constituting a seasonally significant proportion of the primary consumer biomass. As insectivorous adults, they may participate in either aquatic or terrestrial trophic assemblages, or both. Most reptiles, which generally have fewer offspring and lower mortality than amphibians, are insectivorous or carnivorous and are more significant as secondary or tertiary consumers in food webs. Although reptiles interact primarily in the terrestrial environment, some species, including turtles (which may be partially or entirely herbivorous) and snakes such as Thamnophis, may feed in aquatic situations.
Two factors make amphibians and reptiles especially important elements in trophic relation systems in Alberta and elsewhere. First, despite the fact that they are rarely seen, these animals may constitute a huge biomass. In a variety of habitats throughout North America, the total biomass of amphibians per unit area has been shown to exceed that of birds and mammals combined. In Alberta, this may well be true of many of the more mesic (moist) habitats, where every year billions of frog eggs are laid. Reptiles, though less fecund, may also be extremely numerous.
This may be demonstrated by reference to the dens of Thamnophis, where thousands of individuals may congregate to overwinter. Secondly, because they are poikilothermic (see Chapter 8), amphibians and reptiles require far less energy to grow and reproduce than do birds or mammals of equivalent size. Thus, relative to other terrestrial vertebrates, they contribute a disproportionate amount of useable energy to the food web. Food-web maintenance is important in any habitat type. However, disruption of fragile systems—such as deserts, where the number of organisms at any trophic level can be relatively small—may be particularly devastating. The richest herpetofauna in Alberta occurs in the semi-arid southeast, which is in just such a delicate balance. For this reason, the recent decline of certain Alberta species such as the hog-nosed snake and northern leopard frog, is especially disturbing.
The size and general demography of amphibian and reptile populations in Alberta are not well known. In general, populations are dispersed for most of their activity periods and only congregate, if at all, for overwintering or short periods of breeding. This aspect of the herpetofauna makes it especially difficult to study characteristics of population ecology. From what is known from other regions of North America, it would seem that individuals have rather low vagility (capability of dispersing) and the geographic area covered by a population is probably quite small for most species.
Gene flow is therefore restricted, and total population size is modest. The extent of genetic exchange between adjacent populations is unknown, but in the case of species such as the short-horned lizard, which occupies a discontinuous habitat, it may be minimal. Many species exhibit periods of "programmed" dispersal which may serve to ensure some genetic interchange between populations, but the extent of this phenomenon among Alberta amphibians and reptiles is unknown. Such dispersals may take place in the juvenile stage or, as in some turtles, may be carried out by females in search of egg-laying sites. The Alberta taxa are typically widely distributed throughout central and / or western North America, suggesting that gene flow sufficient to maintain species cohesiveness does occur.
However, the populations in our province are often at the northern edge of their range (only Rana sylvatica extends much further north and only Bufo hemiophrys, the Canadian toad, is centred around our latitudinal band) and run the risk of losing contact with the main body of the species and becoming peripheral isolates. This may have occurred in the case of the Canadian toad, which has close affinities to Bufo america- nus, a species typical of eastern North America. It may have speciated quite recently in association with environmental changes and geographic isolation brought about by glacial movements.
For any species, the ability of individuals to recognize conspecifics as potential mates is essential to the maintenance of cohesiveness. Certain features of general ecology, such as habitat preference and activity pattern, serve to segregate species and maximize the chance of mate encounter. Beyond this, however, specific behavioural traits allow specific recognition to be established. In salamanders, such as Ambystoma, fertilization is internal, and a complex pattern of species- specific courtship movements must be performed by the male in order to entice the female to accept his spermatophore. Pheromones (hormones that act between individuals) also play a role in salamander courtship, especially in those forms with terrestrial mating.
In frogs, specific patterns of vocalization serve as the primary premating isolating mechanisms. Females respond preferentially to the call of conspecific males and may even gather information about age or size, and thus fitness, from these mating calls. Once in proximity, females are grasped by males and held in amplexus (pectoral or pelvic embrace that ensures correct placement of the male and female cloacae for fertilization to occur). In frogs and many other amphibians and reptiles, there may be pronounced sexual dimorphism, with either males or females achieving greater size (Fig. 7.2).
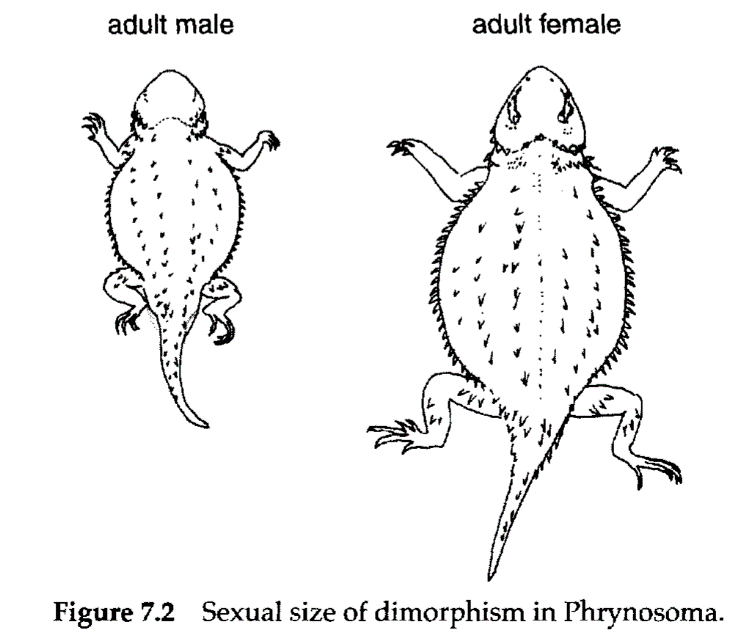
Large female size is often attributed to the greater reproductive output of this sex and the additional space required for gamete production and egg/embryo storage. Larger male size is often related to sexual selection and predominates in species that are territorial or in which competition for mates occurs. Dimorphism may also relate to non-reproductive parameters, such as differences in the diet of males and females. (See species accounts in Chapter 5 for information about dimorphism in Alberta species.)
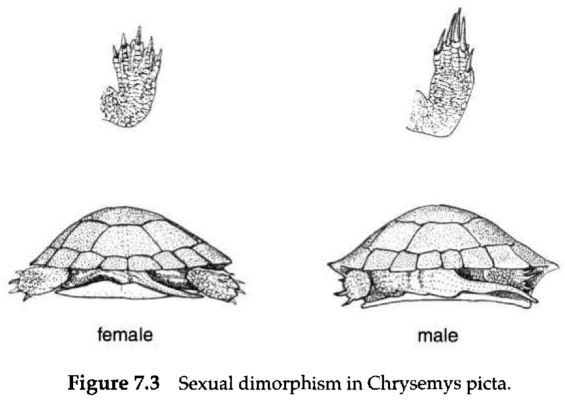
Fertilization in reptiles is always internal and usually involves copulation. As in salamanders, reproduction in reptiles also involves complex behaviours that maximize the likelihood that only intraspecific matings will occur. Chrysemys picta males use their elongate claws in a courtship ritual (Fig. 7.3) and most snakes employ species-specific stereotypical behaviour in conjunction with pheromonal communication in mating. All terrestrial reptiles in Alberta congregate in winter dens or hibernacula. Mating usually occurs early in the active season, just as animals are leaving their winter dens.
This timing takes advantage of the proximity of individuals and removes the difficulty faced by solitary individuals in locating a mate. Reptiles at dens and amphibians at ponds have high site- fidelity and are able to use a variety of biotic and abiotic clues to locate these gathering places year after year. Females of some species, including Thamnophis sirtalis in Alberta, are able to store sperm and delay fertilization—in the event of a season in which no mating is achieved, successful reproduction could still occur.
The ecology of the Alberta herpetofauna cannot be fully under- stood without giving special consideration to the complex life history of amphibians. Larval amphibians constitute a truly distinctive life form. Their biology is almost wholly separate from that of the adults. Larval amphibians cannot be described as "poor tetra- pods/' nor likened to fish. Like all larvae, they exist to feed, grow, and accrue the energetic reserves necessary for metamorphosis into the adult form. Most tadpoles (and some salamander larvae) have relatively short pre-metamorphic lives which correspond to a period of high algal or prey productivity.
Thus, in Alberta, the warm, rainy early summer is the prime time to find most larval amphibians. The necessity to match the larval period with peak productivity accounts for the early breeding of many species, in some cases even as the snow is beginning to melt. All Alberta amphibians lay eggs in an aquatic environment. These vary from species to species in size, shape and type of gelatinous coating. The eggs themselves may be laid singly or in strings or clusters (see Chapter 5). Because they are not guarded by the parents, eggs run a greater risk of predation and destruction due to environmental fluctuations. Pigmentation of the embryo or its coating may shield it from excessive solar radiation.
The shape of the egg mass may promote gas exchange, or in some cases, symbiotic algae resident in the egg may provide oxygen to the developing embryo. Hatching is triggered by temperature, pH, and especially oxygen availability. Hypoxia appears to be the proximal signal that initiates the production of enzymes used to break down the vitelline membrane and egg capsule in those species that have been investigated.
Both salamander larvae and frog tadpoles possess large guts, suit- able for the processing of copious amounts of food or filtrate (Fig. 7.4).

Their mouthparts are specialized for ingestion in the dense aquatic medium. Typically, salamander larvae employ "gape and suck" feeding mechanisms, whereby relatively large prey items are rapidly drawn into the mouth via a complex negative-pressure system. In this type of feeding, a column of water containing the prey is drawn into the pharynx, where gill rakers strain out the solid material, allowing the fluid to pass out of the body via the gill slits. Frog tadpoles tend to employ less spectacular feeding modes. Some types filter large quantities of detritus across their gills, removing tiny particles of digestible material.
This is a rather indiscriminate approach to nutrition, with the animal selecting out valuable items only after ingestion. Water taken in with the food is expelled through the spiracle, the common opening of the internal gill chambers. Other tadpoles feed on algae or carrion which is scraped away by their horny teeth. These teeth are unrelated developmentally and evolutionarily to the true adult teeth. Their patterns are typically complex and are used as key features in the identification of larval specimens (see Chapter 5).
Just as the feeding mechanisms of larvae are related to the aquatic medium, so are their modes of locomotion. The chief device for producing thrust in swimming is the tail. In salamander larvae, both the tail and body are thrown into S-shaped undulatory waves which pass posteriorly, generating thrust as the tail fins and lateral surfaces of the animal are pushed against the water. The globular body of frog tadpoles, however, precludes its incorporation into the generation of propulsive waves. Rather, the tail alone sweeps or sculls to move the animal. Salamander larvae, which develop limbs relatively early in ontogeny, are also able to walk on the substrate, but rely on lateral undulation for any rapid locomotion.
All larvae will attempt to swim away from danger, but their speeds are slow when compared to those of many predators, which include fish, aquatic insects, snakes and other amphibians. Many forms have evolved noxious or even poisonous skin toxins to deter predators (see Chapter 10). At least some tadpoles are able to warn their siblings, and in some cases other species, of danger. Larvae that have been wounded by a predator release "Schreckstoff," a chemical alarm substance, into the water. Its effect is to cause the other members of the school to flee.
Chemical communication, which is highly effective in water, is also used to regulate coordinated schooling behaviour in tadpoles. Amphibian larvae also obtain information about their surroundings via the lateral line system, a series of mechanoreceptors sensitive to vibration and angular displacement. The system may also have electroreceptive capabilities useful in detecting prey. These receptors are similar in construction to the receptors of the inner ear of all vertebrates. During metamorphosis the lateral line system degenerates, as it cannot function in air, and is therefore unnecessary for the chiefly terrestrial adult amphibian.
Date added: 2022-12-11; views: 1359;