Control of Secretion of Adrenal Gland Hormones
A. Adrenal Cortex. Cortisol is secreted by the adrenal cortex in response to ACTH, a peptide hormone secreted by the human pituitary gland. ACTH secretion occurs in response to decreased circulating concentrations of cortisol, as part of a compensatory negative feedback system, and in response to stressors of many types, including surgery, hemorrhage, thermal injury, and hypoglycemia (reduced concentrations of glucose in the blood). In addition, there is a circadian (daily) rhythm of ACTH and cortisol secretion, which results in increased production toward the end of the sleep period and, thus, higher levels of circulating cortisol in the morning than at night.
ACTH is a 39-amino acid peptide derived from the larger molecule pro-opiomelanocortin (POMC), which is secreted under the control of neurotransmitters and corticotropin-releasing hormone, a 41- amino acid peptide of hypothalamic origin. The relationship of ACTH to POMC is shown in Fig. 3. While there is considerable interspecies variability of POMC structure in many parts of the molecule, the first [N-terminal] 24 amino acids of ACTH are highly conserved among species and are biologically active. Evidence from animal experiments indicates that non-ACTH POMC peptides may synergize with ACTH in control of glucocorticoid secretion.
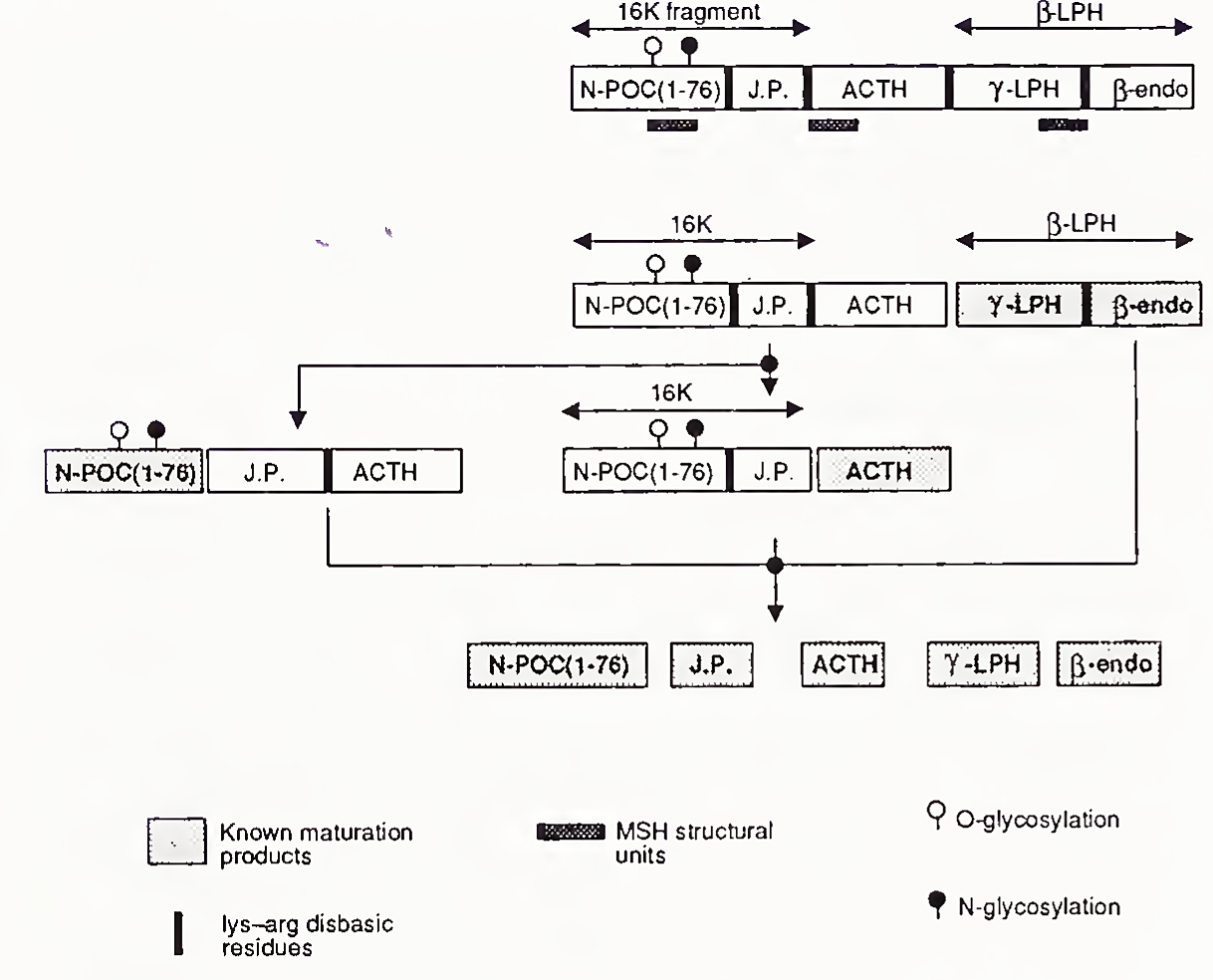
FIGURE 3. Structure and processing of pro-opiomelanocortin (POMC, Poe), endo, endorphin; J.P., joining peptide; LPH, lipotropin; MSH, melanocyte stimulating hormone. [Reproduced, with permission, from N. Seidah, J. Rochemont, J. Hamelin, S. Benjannet, and M. Chreitien (1981). The missing fragment of the pro-sequence of human POMC; Sequence and evidence for C- terminal amidation
ACTH binds to specific adrenal cell membrane receptors, activates adenyl cyclase, and thereby causes an increase in intracellular concentration of cyclic 3',5'-monophosphate (cAMP). This in turn causes an increase in cellular protein kinase activity (which adds phosphate groups to proteins), and an increase in activity of cholesterol ester hydrolase. This enzyme produces free cholesterol needed for the synthesis of pregnenolone, a precursor of steroid hormones. Plasma lipoproteins also provide cholesterol for steroidogenesis.
As shown in Fig. 4, pregnenolone can be converted to mineralocorticoids, glucocorticoids, or androgens. Many of the microsomal enzymatic steps in this conversion are controlled by ACTH, which regulates the synthesis of the enzymes.
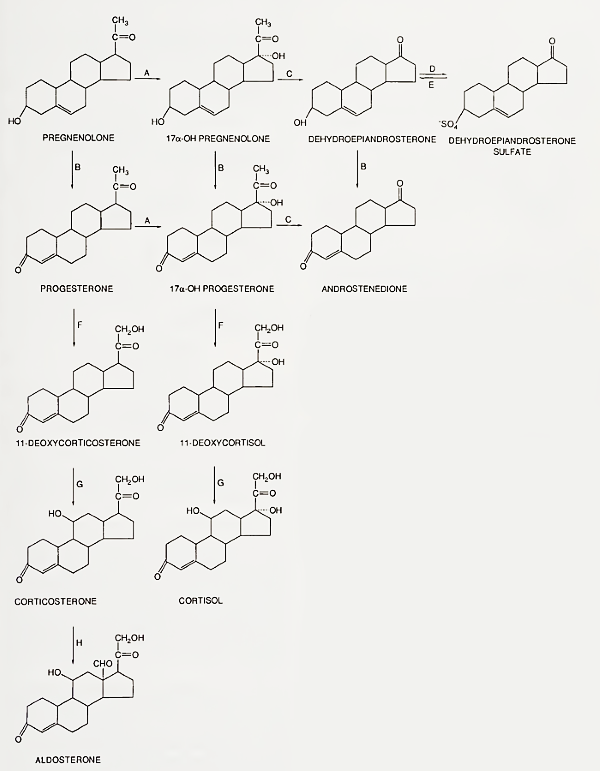
FIGURE 4. Human adrenocortical steroidogenic pathways. Enzymes: A, 17-hydroxylase; B, 3/3-hydroxysteroid dehydrogenase-isomerase; C, C17-20-desmolase; D. steroid sulfotrans-ferase; E, steroid sulfatase; F, 21-hydroxylase; G, 11-hydroxylase; H. 18-hydroxylase, 18-dehydrogenase
Adrenal androgens are also secreted in response to acute ACTH stimulation, but their control is more complex because in some situations they are not secreted in conjunction with cortisol. These situations include adrenarche, puberty, aging, polycystic ovarian syndrome, stress, and starvation. Adrenarche is the process of adrenal gland maturation, which occurs before puberty at approximately 7 yr of age, that involves increased secretion of DHA and DHAS with constant secretion of cortisol. Throughout life, while basal levels of cortisol are unchanged (Fig. 5), basal concentrations of DHAS undergo marked variation, with a peak in the third decade, and a sharp decline thereafter.
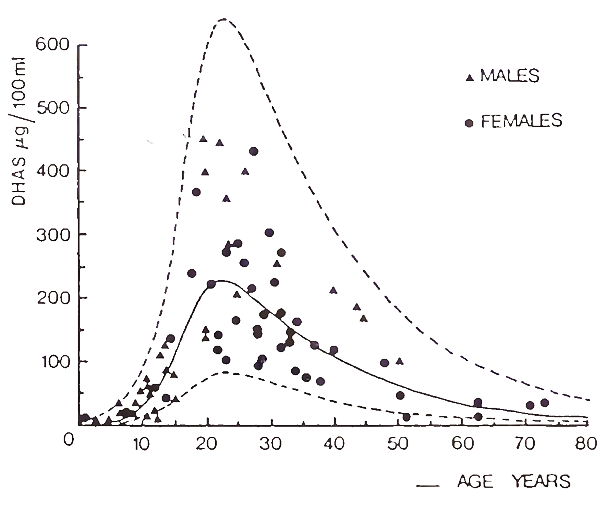
FIGURE 5. Serum concentrations of DHAS in normal subjects, 1-73 yr of age
Aldosterone is produced only by the zona glo- merulosa, the only zone with the necessary enzymes (18-hydroxylase and 18-dehydrogenase). The zona glomerulosa does not produce cortisol because it lacks the enzyme 17-hydroxylase. Aldosterone secretion is controlled by several factors including ACTH and angiotensin II. Angiotensin II production is controlled by renin, which mediates conversion of a hepatic precursor (angiotensinogen) present in the blood to the 10-amino acid peptide angiotensin I; this, in turn, is converted to the 8- amino acid peptide angiotensin II by a converting enzyme in lung and other tissues (Fig. 6).
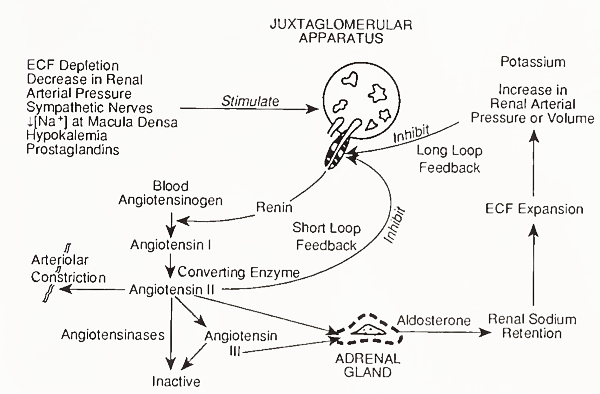
FIGURE 6. Control of aldosterone secretion by the renin-angiotensin system. ECF, extracellular fluid
Renin release is controlled primarily by the sodium concentration of fluid in contact with the renal juxtaglomerular cells and by the renal blood pressure, as sensed by renal baroceptors. Increased renin release is caused by decreased total body sodium or blood pressure. The 7-amino acid peptide angiotensin III is also bioactive.
Angiotensins II and III bind to receptors in zona glomerulosa cells and stimulate aldosterone secretion by a calcium-dependent mechanism involving activation of protein kinase C, which is independent of cAMP. The action of angiotensins may be mediated by prostaglandins, especially of the E series, and inhibitors of prostaglandin synthesis inhibit the effects of angiotensin II on aldosterone secretion.
Potassium ions also influence aldosterone secretion. An increase in serum potassium ion concentration of 1 meq/liter may triple the rate of aldosterone secretion. This is a direct effect on zona glomerulosa cells and forms the basis for a feedback mechanism that regulates the concentration of extracellular potassium ions. Concentrations of potassium ion have the opposite effect on renin concentrations, but the direct effect on aldosterone secretion is predominant. In addition, as in the case of cortisol and adrenal androgen secretion, additional control of aldosterone secretion may be exerted by non-ACTH POMC-related peptides.
B. Adrenal Medulla. Control of secretion of the adrenal medulla is best understood by considering the functional organization of the sympathetic nervous system. Whereas preganglionic fibers of the parasympathetic branch of the autonomic nervous system emerge from cranial and sacral spinal nerves, those of the sympathetic nervous system emerge from thoracic and lumbar spinal nerves and innervate many organs, including the adrenal medulla, as shown in Color Plate 1. These fibers then terminate in ganglia of the paraspinal sympathetic trunk, nearby plexuses, or in the adrenal medulla.
Preganglionic nerve impulses are transmitted to the ganglia neurons by liberation of acetylcholine at nerve terminals. This results in secretion of catecholamines by the peripheral sympathetic nervous system and by the adrenal medulla. Norepinephrine is the major secretory product of the peripheral nervous system, whereas in the human adrenal medulla, the ratio of epinephrine to norepinephrine secretion is approximately 4:1.
Catecholamine biosynthetic pathways are shown in Fig. 7. As in the case of steroids, specific enzymes catalyze synthetic steps. The rate-limiting step in catecholamine biosynthesis is the initial conversion of tyrosine to DOPA by tyrosine hydroxylase. Tyrosine itself is derived from the diet or converted in the liver from phenylalanine by phenylalanine hydroxylase. Dopamine is also found in sympathetic neurons and the adrenal medulla.
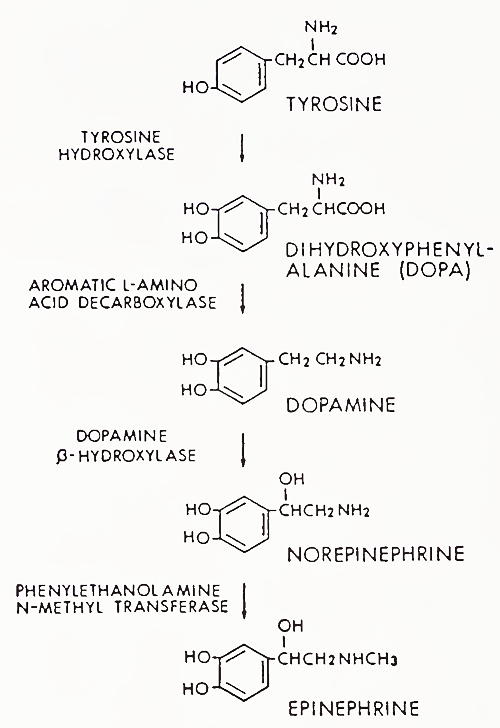
FIGURE 7. Catecholamine biosynthetic pathway of the sympathetic nervous system
The major difference between the pathways of the adrenal medulla and the peripheral sympathetic nervous system is that the medulla contains the enzyme phenylethanolamine-Af-methyl transferase (PNMT). Synthesis of this enzyme is induced by high concentrations of cortisol present in the (portal) circulation from the adrenal cortex to the medulla.
A large percentage of catecholamines released by neurons at nerve terminals are inactivated by reuptake into storage granules of the terminal. Metabolism of circulating catecholamines occurs via two main pathways, mediated by the enzymes catechol- O-methyl transferase (COMT) and monoamine oxidase (MAO), as shown in Fig. 8. The end product of norepinephrine and epinephrine metabolism after conversion by both enzymes is 3-methoxy- 4-hydroxy-mandelic acid (vanillylmandelic acid [VMA]).
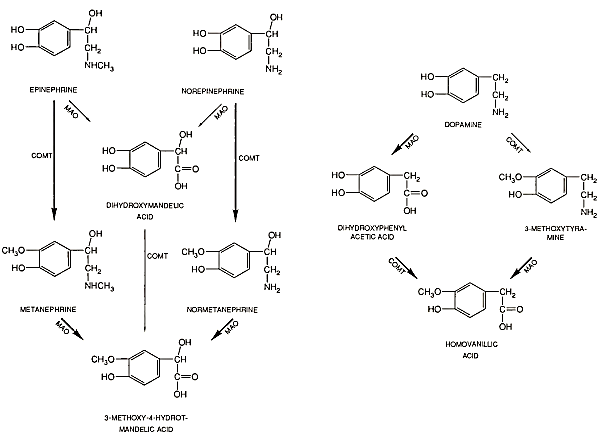
FIGURE 8. Metabolism of catecholamines by catechol-O-methyltransferase (COMT) and monoamine oxidase (MAO)
The hypothalamus is the main regulator of sympathetic nervous system function. Impulses from the posterior and lateral hypothalamus result in generalized discharge of the sympathetic nervous system, including the adrenal medulla. As discussed above, this discharge occurs in response to a variety of noxious, threatening, or stressful stimuli.
In addition, the sympathetic nervous system is instrumental in maintaining an appropriate blood volume during changes of posture from supine to upright, by regulating pulse rate, cardiac output and blood vessel resistance. These feedback systems are mediated by sensors in the carotid sinuses, aorta, and medulla, which detect changes in circulatory volume and blood pressure.
Date added: 2023-05-09; views: 1023;