Avoidance of Light Stress and Permanent or Dynamic Acclimation
Lack of light, interspersed with high light intensity of short-term sunflecks, combined with a change in the spectral composition of the radiation (Fig. 3.2), is a major stressor of the vegetation on the forest floor and, less dramatically, in the shade crown of a tree. As the leaves in the canopy absorb mainly blue and red light, green light is enriched in the spectral composition of the subcanopy space (“green shade”). Not visible to the human eye is another change in the spectrum of sunlight by passage through the forest canopy: the dramatic decrease in the red to far-red ratio, as the leaves absorb red and blue light but transmit and reflect far-red light.
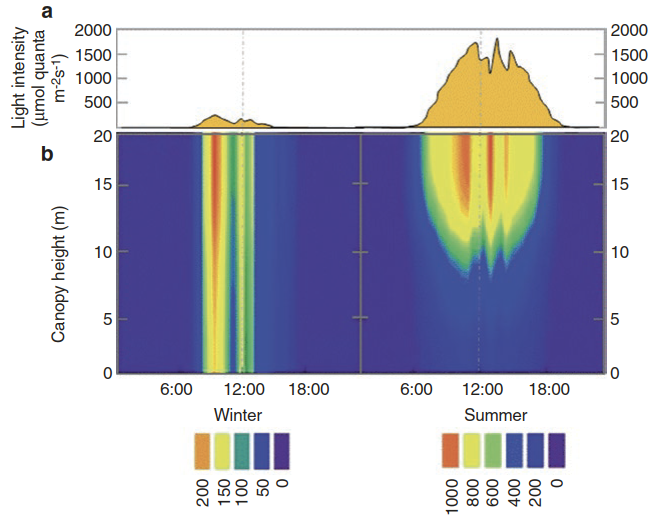
Fig. 3.2. Seasonal change of the light climate in an oak-pine forest. a Daily courses of the light intensity above the canopy. b Light attenuation in the New Jersey Pine Barrens oak-pine forest in early February (left side) and at the end of July (right side). Note the different scales used for the colour bars for winter and summer. All units are in micromoles of photons per square metre per second
Responses to low light are obvious at the forest edges where subcanopy plants (e.g. ferns and other herbaceous species) show a pronounced phototropic reaction—that is, they grow towards the higher radiation intensity outside the forest. Plant life forms that avoid the attenuated light climate under the canopy of a forest are functional types such as epiphytes and lianas; both are light parasites that use trees as support to reach more favourable light conditions.
On the other hand, a surplus of light intensity can also cause problems, as photosynthetic pigments cannot avoid energy absorption, which may overstrain the photosynthetic capacity of the chloroplasts and produce reactive oxygen species (ROS). Leaves often avoid excessive irradiation by adopting a parallel position to the incident light. The hanging leaves of Eucalyptus are well known from the “shadeless forests” in Australia. Likewise, the upright position of the leaves of the characteristic giant rosette plants of the tropical high mountains (Fig. 4.25) has been described as a mechanism for avoiding direct solar radiation.
The position of the leaf lamina of many plants varies during the daily light period, changing the angle and intensity of the incoming radiation. Such leaves often have joints between the petiole and the lamina, known as pulvini (Fabaceae, Oxalidaceae), which enable fast changes in the position of the entire leaves or at least the pinnules. The reaction of the North American wood sorrel (Oxalis oregano) to shortterm high-light stress from a sunfleck is shown in Fig. 3.3.
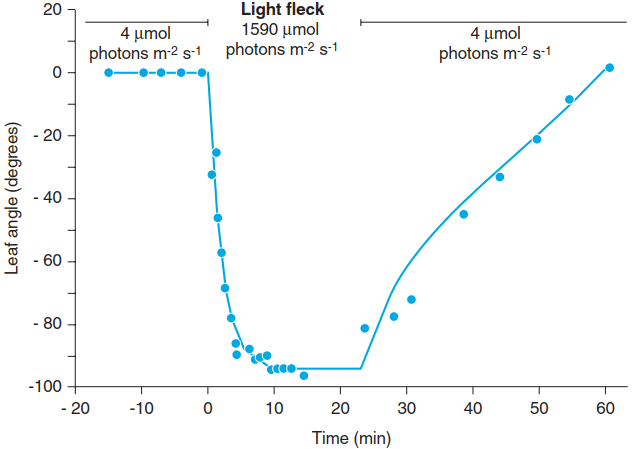
Fig. 3.3. Avoidance of Light stress by a change in the Leaf angle. Oxalis oregano, the wood sorrel, is a particular shade pLant whose leaves avoid light stress of bright sunflecks by rapidly changing the angle of their leaflets by virtue of pulvini. When the sunfleck is over they slowly return to their horizontal position, which is optimal for light harvesting. (After Bjorkman and Powles
Dynamic acclimation to the light intensity can be observed at the cellular level too. In many algae and mosses, but also in vascular plants, chloroplasts can change their position in the cell. They accumulate on the light-exposed surfaces in the case of low light and at the light-parallel cell boundaries when the radiation becomes too strong (Fig. 3.4). Such readily reversible intracellular reactions are mediated through the blue light receptor phototropin (Wada 2013).
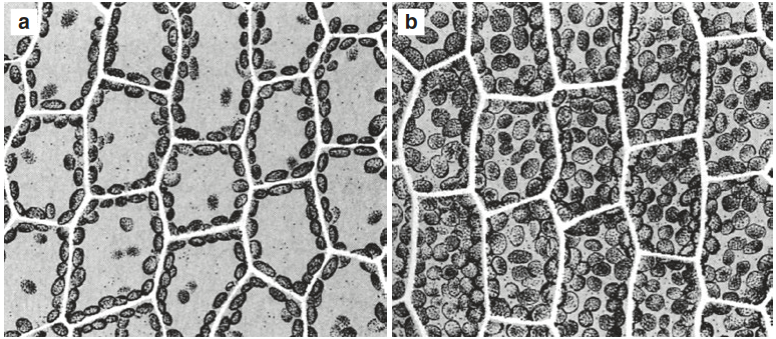
Fig. 3.4. Chloroplast movement is important for avoiding photodamage under high Light and for efficient Light harvesting under Low Light. Object: Leaflet of the moss Funaria hygrometrica, consisting of a single Layer of chloroplast- containing cells. a Position on the lateral walls of the cells in high Light. b Position of the chloroplast on the Light-exposed side in Low Light (Nultsch 2001). A special protein named Chloroplast Unusual Position 1 (CHUP 1) is unique in positioning and moving chloroplasts in the Arabidopsis thaliana leaf cell. It is assumed that CHUP 1 connects the chloroplast with the actin cell skeleton and the cell membrane
Besides such short-term acclimations (with a reaction time in minutes), development of leaves responds to the light environment by the so-called shade avoidance syndrome and the formation of sun and shade leaves. The shade avoidance syndrome is part of the morphogenetic reaction of seedlings when they are grown under low light intensities caused, for instance, by competing neighbours. Owing mostly to the identification of the main photoreceptors in the model plant Arabidopsis thaliana and the availability of respective mutants, knowledge on the molecular regulation of plant growth and development by light is rapidly accumulating Because a separation of acclimation and development is principally difficult here, we focus on the reactions triggered by light stress—that is, by either insufficient or supraoptimal light intensities.
The typical shade avoidance reaction in A. thaliana can serve as an example illustrating the integration of light cues and growth at the molecular level. Seedlings show an inhibition of lamina expansion, while petiole elongation and stem growth are enhanced. Phytochromes (PhyB) and cryptochromes (Cry1 and Cry2), as photoreceptors, have been demonstrated to play specific roles in these differential growth responses. Phytochromes especially are ideally suited to convey information on shade occurrence because of their switching between red and far-red absorption and the pronounced difference in the ratio of these two light qualities between sunlight and shade light. Low red to far-red ratios typical of shade light reduce the levels of active PhyB.
Because PhyB inactivates a class of transcription factors named phytochrome-interacting factors (PIFs), their activity increases under shade light conditions. PIFs have been demonstrated to induce growth responses—for instance, through the activation of genes involved in cell wall biosynthesis. In addition, PIFs influence phytohormone signalling. They enhance auxin concentrations through the activation of auxin biosynthesis. Promotion of shade avoidance reactions by the other growth hormones gibberellic acid and brassinosteroids can be attributed to a positive effect on PIF abundance. The function of cryptochromes in shade avoidance is associated with degradation of the constitutive photomorphogenesis protein 1 (COP1) (Fig. 3.22), which is jointly triggered by cryptochromes and phytochromes, albeit in an unknown fashion different from photomorphogenesis (Casal 2013).
The shade avoidance reaction as observed under low red to far-red illumination must be attenuated when exposure to this light climate is prolonged and also when the leaf reaches into a space with sufficient light intensity. Similarly, sun- flecks reduce the shade avoidance response. An autoregulatory mechanism involving transcription factors such as HY5 (Figs. 3.21 and 3.22) has been described that helps to avoid exaggerated shade avoidance responses (Jiao et al. 2007).
Through the shade avoidance reaction the plant strives to escape unfavourable low light intensity, using its resources to grow into a better-illuminated space. A typical example is the ground flora in a forest, consisting of herbs and bushes with elongated internodes and smaller leaves. The formation of sun and shade leaves, on the other hand—which is well known from tree crowns—can be understood as differential acclimation of a plant’s foliage to high light intensities in the outer part (the so-called sun crown) and low light intensity in the centre of the crown (termed the shade crown).
Such differentiation within the same shoot is possible because of the modularity of the plant structure, which is composed principally of similar units called phytomers, whose development can be individually guided by local environmental cues. Sun leaves deliver the major share of the photosynthetic gain owing to an acclimated photosynthetic machinery but also exhibit greater dynamics of cell respiration. In shade leaves, both processes run at lower rates, which shows their autonomy and ensures survival. Export of assimilates from shade leaves to other plant organs, however, may be low or non-existent. Further characteristics of thick sun leaves and thin shade leaves are summarised in Table 3.1.
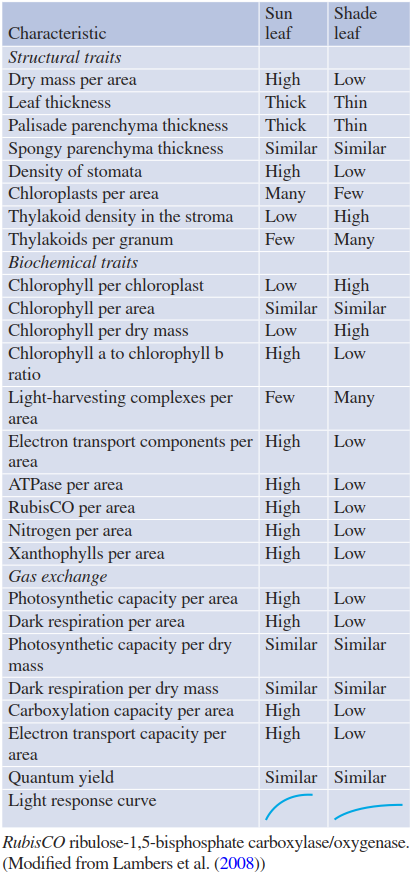
Table 3.1. Comparison of characteristic traits of shade-and sun-acclimated leaves
In contrast to trees, herbs are amenable to experimental approaches aimed at understanding the formation of physiologically and anatomically different types of leaves (Fig. 3.5). The signals indicating a high-light climate are perceived by the mature leaves, which trigger the respective development of the young leaves in the apical shoot meristem via long-distance signalling. Interestingly, the ambient CO2 concentration has effects on the formation of sun and shade leaves similar to those of the light intensity. High CO2 leads to the formation of sun leaves (Prior et al. 2004; Pritchard et al. 1999).
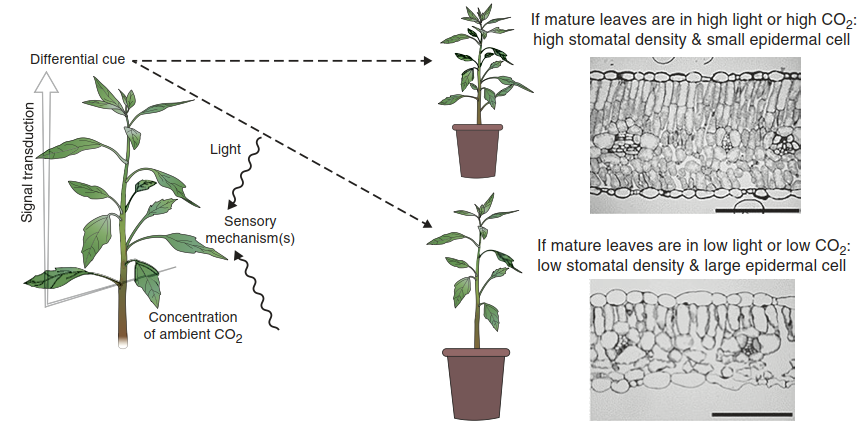
Fig. 3.5. Differential development of sun and shade leaves in Chenopodium album in response to light intensity and CO2 concentration. Sun leaves have a dense mesophyll with at least two layers of palisade parenchyma cells, which originate from a directional change of the cell divisions. The scale bars indicate 100 μm
From experiments such as those shown in Fig. 3.5 it was concluded that, at least in dicots, systemic signaling depending on the light environment influences the development of new foliage and thus triggers acclimation to the actual light quality and intensity. Long-distance signalling from mature to developing leaves is also known from herbivore and pathogen attacks, and most likely a multitude of signals is involved in mediating different types of responses. Acclimation of the photosynthetic apparatus appears to be under the control of the redox state. Thus, redox signalling may also be part of the long-distance conveyance of biochemical information. Also, the concentrations of photosynthates may act as an indirect cue for the light climate of mature leaves. However, since the formation of sun and shade leaves is a highly complex syndrome, involvement of various further signals can be expected.
At the cellular level the epidermal cells, in particular, contribute to the protection of the mesophyll cells against high-light stress. A felt-like layer of dense hairs, which reflects incident light, is found on young leaves of many plant species. Although it can be kept for permanent protection, it mostly disappears during maturation of the leaves. Protective pigments, which absorb predominantly shortwave radiation, are also found in the epidermis of young leaves. A typical example is the so-called juvenile anthocyanin (Fig. 3.25), which protects not yet fully green leaves during the development of the photosynthetic apparatus. Later, protection is conferred mostly by carotenoids in the antennae. This is apparent from the disappearance of the pigments in the epidermis.
Date added: 2025-01-13; views: 524;