Inhibition of Shoot Growth. Stimulation of Root Growth
Shoot growth responds very rapidly to water deficit. Acclimative slowing of growth can be observed within 20-30 min. The response occurs even in the absence of any changes in the water potential of the elongating cells. Thus, it is not a mere consequence of turgor loss. Furthermore, growth halts much faster than photosynthesis under drought conditions. A shortage of reduced carbon can therefore be ruled out as the cause of the growth reduction as well. In fact, sugars often accumulate after the onset of stress, meaning that growth is uncoupled from the availability of carbon under these conditions, once more indicating controlled processes rather than simple supply- driven processes.
An important inference from active growth modulation is that growing tissues and mature tissues—for example, young sink leaves and old source leaves—respond differentially to stress (Fig. 6.16). This principal difference is indicated by many observations but has not been systematically dissected yet at the molecular level (Claeys and Inze 2013).
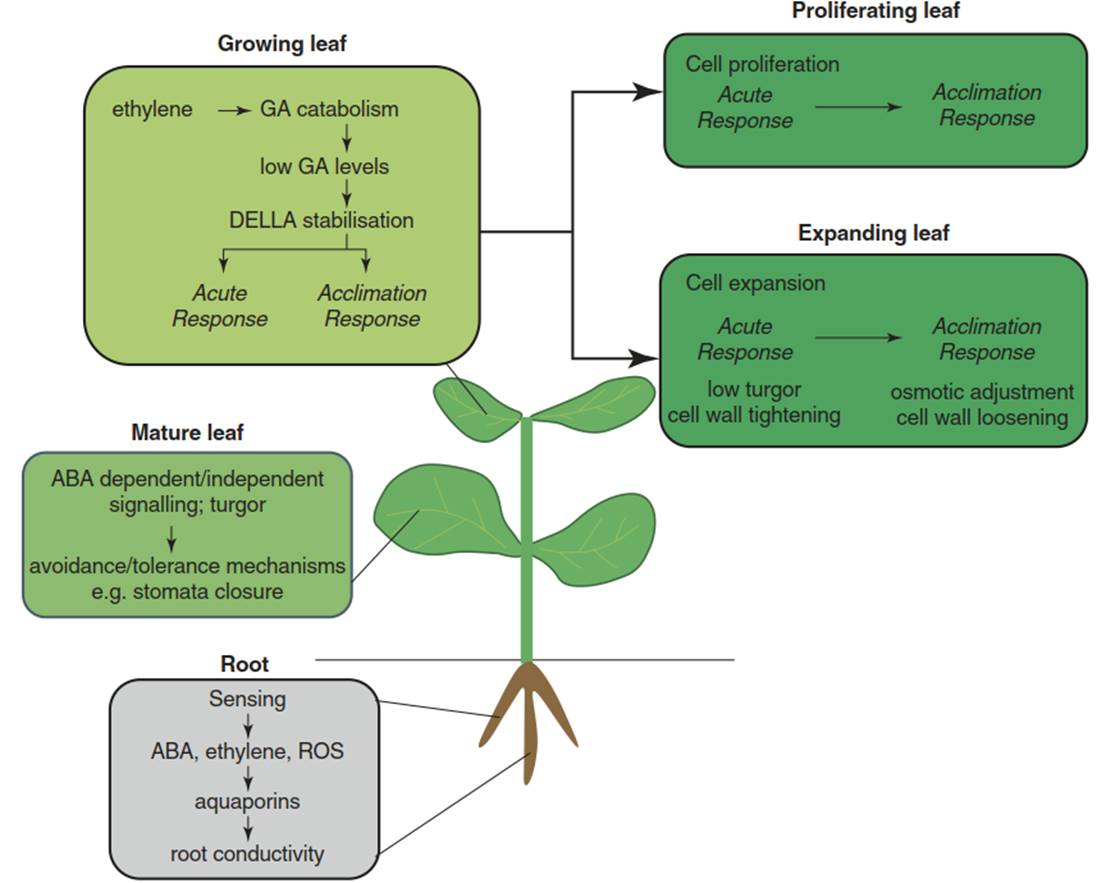
Fig. 6.16. Processes involved in growth regulation upon drought. The responses differ between mature and growing leaves and, in the latter, between proliferating leaves (growth by cell division) and expanding leaves (growth by cell expansion). Soil drying is sensed in roots, activating a combination of hydraulic and chemical signals, which are transported through the xylem to the leaves (Fig. 6.18), where they initiate a number of tolerance mechanisms.
The response of mature leaves can be described by the avoidance/tolerance concept. In growing leaves, stress leads to acute growth inhibition followed by growth acclimation, both mediated by DELLA signalling—that is, the inhibition of gibberellic acid signalling. Ethylene promotes DELLA stabilisation and growth inhibition. While hormonal signalling is common between expanding and proliferating leaves, effector genes are distinct. In proliferating leaves, inhibitors of the cell cycle might play a role in the acute response. In expanding leaves, cell wall tightening and changes in cell turgor lead to growth cessation, while osmotic adjustment and cell wall loosening are important for growth acclimation. (Modified from Skirycz and Inze (2010))
Growth is determined by the cell division rate and cell expansion rate. Under water-limited conditions, both processes are actively modulated in shoots. Cell cycle activity, which determines the cell division rate, is triggered by cyclin-dependent kinases and associated cyclins (see molecular biology and biochemistry textbooks). The major pathways controlling cyclin-dependent kinases are all responsive to drought. For instance, repression of the anaphase- promoting complex/cyclosome (APC/C), a master negative regulator of cell cycle activity in eukaryotes, is down-regulated, leading to reduced cyclin-dependent kinase activity.
The cell cycle control pathways themselves are controlled by DELLA proteins as central regulators of gib- berellic acid signalling and growth. The stress hormone ethylene has also been implicated in cell cycle arrest. A very early transcriptional response to osmotic stress specifically in growing leaves is the up-regulation of ethylene response factors that stimulate the catabolism of gibberellin (Claeys and Inze 2013).
Cell expansion requires turgor pressure and sufficient extensibility of the cell wall. Aquaporin-facilitated water uptake can lead to cell enlargement, provided that the osmotic driving force for water uptake—that is, the intracellular concentration of solutes such as K+—is strong enough. The higher the rigidity of the cell (the lower the extensibility of the cell), the higher the pressure required to expand the cell. The dynamics of cell wall characteristics therefore play a central role in plant growth processes.
Cell wall flexibility is modulated, on the one hand, by ROS-dependent cross-linking and cleavage reactions and, on the other hand, by enzymes modifying the structure of pectins or influencing the interaction between cellulose microfibrils and hemicelluloses (for cell wall structure, see plant biochemistry and plant physiology textbooks). Such enzymes include pectin methylesterases, xyloglucan endotransglucosylases/hydrolases and expansins. Their expression patterns change substantially under conditions of drought (Tenhaken 2015).
The precise and high-resolution description of cell wall dynamics and growth processes at the cell, tissue and organ levels is a complex systems biology problem, which is very actively studied and far from being solved. However, three aspects can be singled out to help illustrate the shoot growth responses under conditions of water scarcity. First, osmotic adjustment can maintain the turgor pressure to enable continued growth under stress and may thus have different significance for cells of growing leaves than for those of mature leaves. In the latter, the main purpose of osmotic adjustment is avoidance of water loss. Second, cell expansion with reduced turgor pressure can be achieved by increasing the flexibility of the cell wall.
Third, growth can be effectively stopped by a stiffening of the cell wall. Shoot growth under drought often goes through two different phases. During the acute phase, growth ceases as a consequence of cell wall stiffening. In a second phase of growth acclimation the cell wall becomes more flexible again to support new expansion growth (Skirycz and Inze 2010).
Depending on the severity of stress and on the genetic variation selected for in habitats varying in the extent of water limitation, different processes can be dominant to establish the right balance for the fundamental trade-off between growth and stress tolerance. For instance, a detailed study of A. thaliana responses showed that under mild drought stress the expression of expansins was up-regulated (that is, cell walls became more extensible and growth was maintained) whereas under more severe drought, expansins were down-regulated (Harb et al. 2010) (that is, cell walls were more rigid and growth was halted).
Plants not only reduce the growth of evaporative surfaces when they are water limited; they also lower the density of stomata per epidermal cell and per unit of leaf area to further reduce water loss. In developing leaves the divisions of epidermal cells are controlled in such a way that under drought conditions, fewer stomata arise. This process is under the control of the mature leaves and their perception of water status. ABA is a key molecule also in regulating stomatal development.
Stimulation of Root Growth. The root system architecture belongs to the “hidden half” of plant biology. Plasticity of root growth has, for a long time, not been studied as extensively as the plasticity of shoot growth. This changed only recently, at least partly fuelled by the expectation that major advances in sustainable agriculture may be achievable by breeding for relevant root traits.
A general shift in plant growth under water limitation is the increase in the root to shoot biomass ratio. This applies at least to situations of mild stress. More pronounced drying of the soil inhibits root growth as well, in part because of high soil impedance—that is, an increase in the force needed to penetrate the soil. Overall, the plasticity of the root system is even greater than that of the shoot. For instance, the surface area can vary more widely because of the tremendous influence of root hairs on the total surface. Mechanistically, the modulation of root system architecture in response to soil water is barely understood. Auxin gradients and cytokinins play a major role in controlling root morphology. ABA is known to influence the respective pathways.
Date added: 2025-01-18; views: 527;