Recognition of Herbivore Attack
Rapid induction of direct and indirect herbivore defences is of course dependent on reliable cues perceived by the plant. Insect feeding causes mechanical damage, and plants indeed respond to this (wounding response). However, detailed comparative studies—for instance, of transcriptome changes—have clearly shown that a plant differentiates between the tissue loss due to mechanical wounding and the tissue loss caused by a feeding insect. The pattern of gene induction and repression is clearly distinct for the two conditions (Howe and Jander 2008).
Thus, there are apparently signals associated with the insect’s activity that provide information for the plant. Sources of such signals can be insect saliva, regurgitant (another form of oral secretions produced by lepidopteran larvae), oviposition fluids or faeces. In analogy to the term MAMPs used in the context of pathogen defence, these signals are now referred to as HAMPs (herbivory associated molecular patterns) or insect elicitors. In addition, the more general term DAMPs is used as well, summarising signals arising from the plant because of enemy activities (e.g. mechanical damage). In fact, the concept of DAMP recognition unifies induced plant immunity to pathogens and herbivores.
The first HAMP identified was volicitin, a fatty acid-amino acid conjugate isolated from oral secretions of beet army worm caterpillars (Spodoptera exigua) (Alborn et al. 1997). The conjugate is derived from a plant fatty acid, while the glutamine and the conjugation reaction stem from the insect. Treatment of artificially wounded maize plants with volicitin activates the production of volatiles typical of the herbivore response, while the wounding alone does not lead to production of the same volatile blend. Since the discovery of volicitin, several other HAMPs have been molecularly identified: a range of other fatty acid-amino acid conjugates, breakdown products of plant adenosine triphosphate (ATP) synthases called inceptins and sulphated fatty acids called caeliferins (Acevedo et al. 2015).
HAMP receptors are not known yet. Generally, recognition of HAMPs by plant cells is molecularly less well understood than MAMP recognition. The same is true of the early signal transduction events triggered by HAMP sensing, even though the existing knowledge shows extensive principal similarity to pathogen responses. The same basic modules are implicated: transient cytosolic Ca2+ spikes, membrane potential changes, ROS as signalling molecules, and mitogen-activated protein kinase cascades (Wu and Baldwin 2010).
As in the plant-pathogen interaction, there is evidence that herbivores produce effectors to overcome plant defences, and that plants have evolved the ability to sense such effectors or their activities. Again, the molecular understanding is far less advanced, at least partly because plant- herbivore interactions are less amenable to the type of genetic analysis that led to the gene-for- gene concept. An example of herbivore effectors are ATPases in insect saliva. Mechanical damage of plant cells causes a release of ATP into the extracellular space. This extracellular ATP can serve as a DAMP for the plant, and insect ATPases degrade this potential signal. Thus, a plant receptor for extracellular ATP presumably functions in DAMP recognition (Choi et al. 2014).
In contrast to the early events of herbivore sensing, the main regulators of plant herbivore responses have been known for a long time.
Jasmonic acid (JA) and its derivatives are recognised as plant hormones that play a central role in balancing a plant’s investments in growth versus defence in rapidly changing environments. JA generally inhibits growth, as well as assimilate export from vegetative tissues, and activates defensive and reproductive processes. Most prominent among the defensive roles of JA is protection from attack by insect herbivores and necrotrophic fungi. JA promotes the synthesis of practically all classes of chemical defence compounds, underpinning its universal role in the plant kingdom. For several plant species it has been shown that a loss of JA synthesis or JA perception severely compromises the ability to limit damage caused by all kinds of insect herbivores and even some vertebrates (Fig. 8.28).

Fig. 8.28. Jasmonate biosynthesis is essential for effective herbivore defence. The tomato defenceless mutant (left)— which is deficient in jasmonate biosynthesis—and wild-type plants (right) were exposed to hornworm larvae for 13 days. The right panel shows the larval size after feeding on the mutant (left) and wild-type plants. (Howe et al. 1996)
Field studies on transgenic N. attenuata plants with a defect in JA synthesis showed far-reaching consequences of JA signalling in the natural environment (Kessler et al. 2004). Not only were specialist herbivores more successful when the capacity of the plants to induce defences was compromised by the silencing of a lipoxygenase gene essential for JA biosynthesis, but also plants were chosen for oviposition and severely damaged by opportunistic herbivores (e.g. Empoasca sp. leafhoppers) that rarely attack N. attenuata plants in the wild. Also, JA-deficient plants were preferred to wild-type plants in choice experiments .
Thus, plants fall victim to herbivores that they normally are able to fend off. Later field experiments revealed that leafhoppers probe plants during initial feeding, and those they select for extended feeding are the ones with the lowest level of JA-dependent activation of defence responses (Kallenbach et al. 2012).
The central position of JA in defence can be described in distinct steps. HAMP or DAMP recognition triggers the early signal transduction chains that activate the synthesis of JA and its derivatives, among them the actual receptor-active JA-isoleucine (JA-Ile) conjugate (Fig. 8.29). JA synthesis proceeds from the ubiquitous fatty acid linolenic acid via enzymatic reactions localised in plastids and peroxisomes (the core JA module). Perception of JA then activates the herbivore defence genes encoding enzymes and regulatory proteins involved in the synthesis of defence metabolites, or the antifeeding defence proteins such as protease inhibitors.
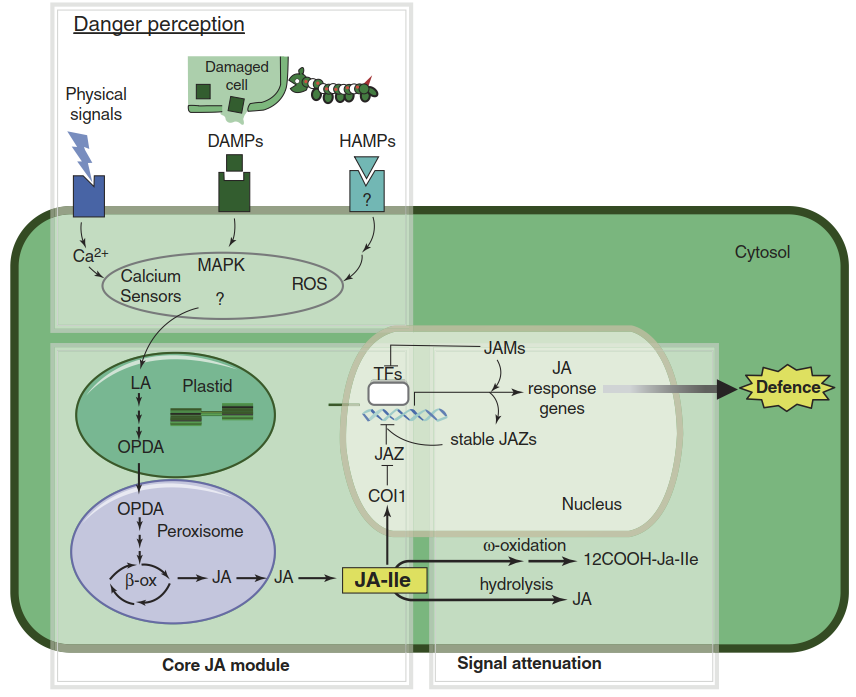
Fig. 8.29. Jasmonate-triggered herbivore defence. Danger signals derived from attacking herbivores (herbivory associated molecular patterns (HAMPs)) and damaged plant cells (danger-associated molecular patterns (DAMPs)) are recognised by pattern recognition receptors (PRRs) at the cell surface. PRR activation triggers intracellular signalling systems involving mitogen-activated protein (MAP) kinase pathways, Ca2+ spikes and reactive oxygen species (ROS) production. In a largely unknown fashion these signalling events activate jasmonic acid (JA) biosynthesis from its precursor, linolenic acid (LA), in plastids and peroxisomes.
JA is the substrate for synthesis of the actual signal JA-isoleucine (JA-Ile) in the cytosol. In the ubiquitin-proteasome system (Fig. 8.30), thereby activating the expression of JA-responsive genes and the expression of defence traits. JA signalling is attenuated by catabolism of JA-Ile via ω-oxidation and hydrolysis, synthesis of JAZ repressors that are stable in the presence of JA-Ile, and transcription factors (JAMs) that negatively regulate JA response genes. OPDA 12-oxo-phytodienoic acid, β-ox β-oxidation, JAM JASMONATE-ASSOCIATED MYC2-LIKE, JAZ Jasmonate Associated Zinc-finger domain, 12-carboxy-JA-Ile 12COOH-JA-Ile. (Modified from Campos et al. (2014))
Upon pathogen or herbivore detection, the active signal JA-Ile, which is barely present in unstressed cells, can be synthesised rapidly (i.e. within minutes) in the cytosol from JA and isoleucine, and then sensed by a receptor in the nucleus (Fig. 8.30). Like several other phytohormone receptors, the JA-Ile receptor COI1 (for “coronatine insensitive”; the name refers to the initial identification of a receptor mutant insensitive to the jasmonate analogue coronatine) is part of an E3 ligase complex that tags specific proteins for degradation in the 26S proteasome by adding ubiquitin chains. These proteins are the JAZ repressors. They prevent the activation of JA-Ile responsive genes by interaction with transcription factors (e.g. MYC2) that bind the promoters of early JA-Ile response genes. Degradation of the JAZ proteins relieves the repression and the JA response is activated (Fig. 8.30).
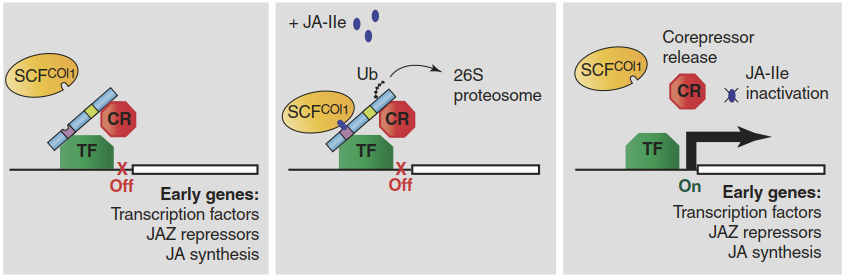
Fig. 8.30. Activation of defence genes upon jasmonic acid-isoleucine (JA-Ile) perception. JAZ proteins (blue bar), supported by co-repressors (CR), bind and repress transcription factors (TF) associated with the promoters of early genes in the JA response. JA-Ile binding to the receptor COI1 promotes interaction of the SC-COI1 E3 ligase complex and the tagging of JAZ for degradation in the proteasome. As a result, early JA response genes can now be transcribed. (Browse 2009)
Herbivore defence responses are analogously switched on in tissues distant from the site of herbivore-inflicted damage. JA or JA conjugates are phloem-mobile signals, which may travel to as yet unaffected tissues. In addition, volatile methyljasmonate serves as an airborne longdistance signal (Fig. 8.26). The JA-dependent activation appears to be universal in the plant kingdom. Besides this central mechanism, there are other known signals and signalling modes. The systemic wound response was discovered and intensively investigated in tomato plants. An early breakthrough was the identification of systemin, a peptide signal derived from a pre-protein through proteolytic processing (Pearce et al. 1991).
Systemin is produced in wounded tomato leaves and activates the JA pathway in neighbouring vascular tissues. It can thus be considered as a DAMP released by wounding. Orthologous peptides with comparable activity have never been found, suggesting that systemin and systemin-like peptides are restricted to a small group of plant species. Nonetheless, defence-activating peptides of a different molecular nature have in the meantime been identified, and they appear to be widespread in the plant kingdom (Bartels and Boller 2015).
A systemic wound response dependent on JA as the mobile signal takes a few hours to materialise. An alternative mode of long-distance signalling could be electrical. Indeed, the up-regulation of JA synthesis in distal tissues within minutes of wounding appears to depend on electrical signalling (Mousavi et al. 2013). Both insect feeding and wounding elicit surface potential changes in A. thaliana leaves, which travel through the rosette to distal leaves at an estimated speed of around 6 cm/min. The amplitude of the signal correlates with the strength of the JA response activation in these leaves. Conversely, the injection of current into leaves systemically triggers JA synthesis in the absence of wounding. Ion channels involved in the generation and propagation of the electrical signal are glutamate-receptor-like proteins.
Date added: 2025-02-01; views: 491;