Inducible Defences Against Herbivores
The previous subchapter provided a glimpse of the huge variety of molecules synthesised by plants that influence the interaction with herbivores, as toxins, deterrents or attractants. While plants produce a complex cocktail of secondary metabolites constitutively, there is, in addition, the induced chemical defence, which comprises not only metabolites but also defence proteins and changes in the storage sites of carbohydrates to minimise the consequences of tissue loss. The advantage of induced defences is that the expenditure of resources is restricted to periods when defence is actually needed, thereby reducing the costs of defence, in favour of growth and reproduction. On the other hand, they have to be activated quickly in order to be effective.
Plant tissues respond locally to herbivore feeding—that is, the cells surrounding the damaged site reprogramme their metabolism and up-regulate the synthesis of defence molecules. In addition, such up-regulation of herbivore defence occurs in tissues distant from the site of attack. As in the case of pathogen attack, longdistance signalling passes the information on herbivore presence to sites not exposed to feeding. The incoming signal (Fig. 8.26) is then converted into changes in gene activities that turn the systemic tissue into a more hostile environment for herbivores. The systemic response to wounding was originally discovered in tomato (Green and Ryan 1972) and later found in a wide range of species.
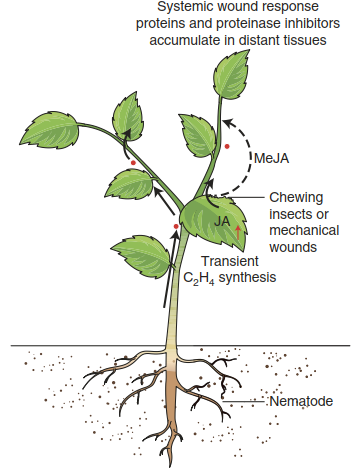
Fig. 8.26. The systemic wound response. MeJA Methyljasmonate
Induced herbivore defences—and, in particular, the systemic responses—add another layer of complexity to interactions within the communities of arthropods, etc., around a plant. Inducible defences connect organisms through space and time. Activities of root-feeding herbivores, for example, influence the chemical composition of above-ground tissues and, with that, the behaviour of organisms responsive to plant volatiles, etc. Similarly, herbivory early in the growing season has long-lasting systemic effects on a plant’s biochemistry and, again, significantly impacts the interacting organisms later in the year.
Plant volatiles emitted by damaged plants can have yet another community consequence. They can result in the so-called priming of neighbouring plants. Perception of airborne signals indicating a herbivore attack on a plant leads to an up-regulation of defence in as yet uninfested plants or to more rapid and robust defence activation when those plants are attacked by herbivores (Fig. 8.27). This has been shown for lima beans (Phaseolus lunatus) exposed to spider mites (Tetranychus urticae). The bean plants released terpenoids, which triggered the activation of defence genes in plants exposed to the volatiles but not to the herbivores (Arimura et al. 2000). Similarly, the treatment of maize plants with volatiles typical of damaged leaves resulted in a much stronger defence response when these plants came in contact with caterpillars (Engelberth et al. 2004).
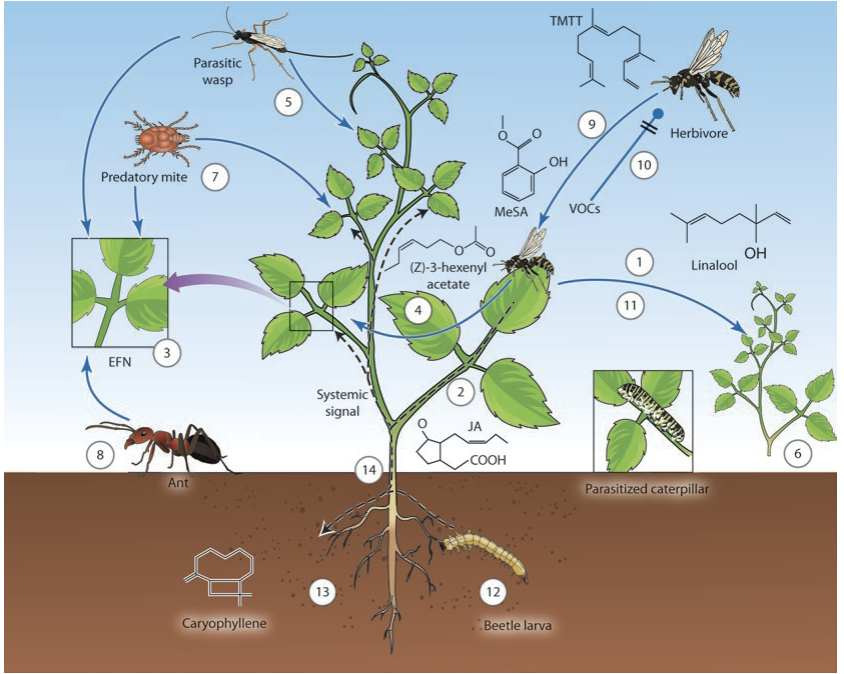
Fig. 8.27. Indirect herbivore defence via a multitude of tritrophic interactions. Biological effects and interactions of volatile organic compounds (VOCs) and extrafloral nectar (EFN). (1) Feeding by a herbivore. (2) Jasmonic acid (JA) synthesis. (3) Inducible release of VOCs and EFN from wounded and intact leaves. (4) Some VOCs can induce indirect defences systemically—that is, in as-yet-unattacked leaves. (5) and (6) VOCs attract parasitic wasps that parasitise herbivores. Attraction of predatory mites (7). Wasps and mites also feed on EFN, as do ants (8). (9) Both con- and heterospecific herbivores can be attracted by VOCs (9), as well as repelled (10). (11) VOCs can also be perceived by other plants of the same species or a different species. (12) Underground feeding by, for example, beetle larvae can induce release of VOCs from roots (13) and elicit a systemic signal triggering EFN production or VOC release from aboveground organs (14). MeSA Methyl salicylate, TMTT 4,8,12,-Trimethyl-1,3,7,11,-tridecatetraene
Most of the defence molecules described above are synthesised throughout the life cycle of a plant. Upon herbivore attack, production of some can be strongly up-regulated, so they are part of both the constitutive and the inducible defences. As a consequence, the attacked plant becomes more toxic and unpalatable. A further step towards this state is the synthesis of proteins that can affect the fitness and growth of herbivores. What is well documented is the production of protease inhibitors that interfere with an insect’s digestive system. A range of different proteins complements the plant defence arsenal.
Oxidative enzymes such as lipoxygenases and polyphenol oxidases have the potential to cause damage to insect tissues. Lectins and other carbohydrate-binding proteins can inhibit sugar-containing molecules in the insect gut. Cysteine proteases damage the peritrophic membrane that protects the gut epithelium. Arginine and threonine deaminases degrade essential amino acids. A feature that many of the plant defence proteins have in common is that they are comparatively resistant to gut proteases. This enables their biochemical identification via selective enrichment during gut passage (Howe and Jander 2008).
Indirect defences can be induced as well (Fig. 8.27). Changes in the blend of volatiles released by a plant upon herbivore feeding are either due to the processing of stored conjugated specialised metabolites (e.g. glucosinolates; Fig. 8.25) or attributable to newly synthesised volatile compounds. A different type of indirect defence relies on rewarding the enemies of herbivores. Secretion of extrafloral nectar attracts non-herbivorous ants and other predators of herbivores (Heil 2008) (Fig. 8.27). The exact chemical composition of such nectars is not known. In an analogy to floral nectaries the presence of sugars such as glucose and fructose, as well as secretion mediated by sugar transporters, can be assumed.
Date added: 2025-02-01; views: 445;