Inducible Systemic Resistance
Besides the cell-autonomous plant immune system there is an additional layer of defence. Cells under attack do not only mount a local response; they also send out signals that activate defence in remote tissues and organs. This phenomenon is called systemic acquired resistance (SAR) (Fig. 8.14). SAR can last for weeks or months and protect a plant from secondary infection—that is, render an otherwise compatible interaction incompatible.
SAR is non-specific in the sense that (1) any potential pathogen (avirulent or virulent) triggers SAR, and (2) protection is achieved against a wide range of pathogens. Furthermore, SAR prepares a plant to respond more rapidly and strongly to subsequent pathogen attack. The plant becomes primed for defence (defence priming).

Fig. 8.14. Systemic acquired resistance of cucumber plants to the anthracnose fungus Colletotrichum lagenarium. One cotyledon of the plant on the left was infected with spores of the fungus. It is suffering severely from necrosis. The plant on the right was left untreated. One week later, 20 drops of a spore suspension were placed on a main leaf of both plants. The leaf of the plant on the left remained fully healthy, as it had developed systemic resistance. The leaf of the plant on the right, which experienced no “immunisation”, exhibited necrotic areas at the positions of every drop of spore suspension
The main changes in systemic tissue involve the synthesis of PR proteins (Fig. 8.15). Unlike ETI, SAR does not involve cell death. Most of the PR proteins are either secreted into the apoplast (i.e. into the space colonised by potential pathogens) or stored in vacuoles. As mentioned above, the actual molecular mechanisms explaining PR protein-mediated protection are barely known. PR protein function is nearly impossible to dissect genetically, because many proteins act in concert and PR genes are often clustered in plant genomes, so multiple knockout mutants are practically impossible to obtain (Fu and Dong 2013).
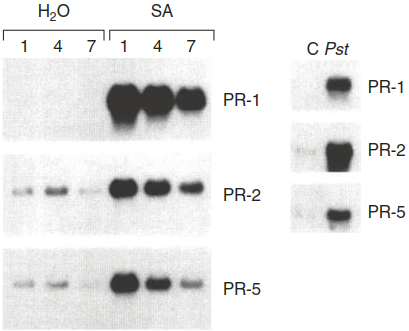
Fig. 8.15. Systemic induction of pathogenesis-related (PR) genes. Treatment of Arabidopsis thaliana leaves with the defence hormone salicylic acid (SA) or inoculation with Pseudomonas syringae strongly activates PR gene (PR-1, PR-2, PR-5) expression. Expression of PR genes was analysed by northern blot hybridisations. C control, Pst Pseudomonas syringae
The trigger for SAR is local ETI—that is, the response of cells under direct attack by a pathogen. ETI leads to an increase in the concentration of the defence hormone salicylic acid. This in turn elicits the generation of phloem-mobile signals, whose molecular nature has not been unequivocally shown (Fig. 8.16). Several compounds are candidates, among them azelaic acid (Jung et al. 2009) and pipecolic acid (Navarova et al. 2012).
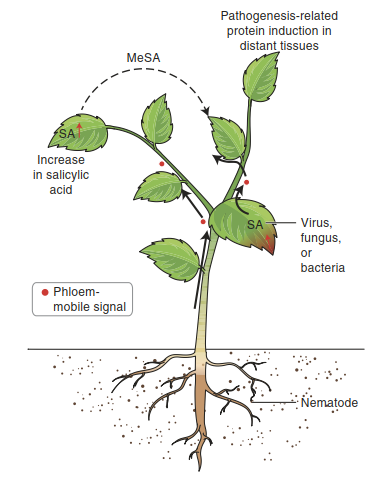
Fig. 8.16. Systemic acquired resistance. MeSA Methylsalicylate
Arrival of the signals in the systemic tissue again results in an increase in the salicylic acid concentration, albeit smaller than that in the local tissue. Salicylic acid is perceived by different receptors locally and systemically. In cells under direct pathogen attack, salicylic acid positively regulates cell death, while in systemic tissue it suppresses cell death and sets signalling cascades in motion that result in PR gene activation and SAR.
Date added: 2025-02-01; views: 632;