Sudden Disturbances and Loss of Pools
Disturbances that result in removal of biomass and resources from the ecosystem (export of resources) have impacts rather different from those disturbances after which pools and thus resources remain in the ecosystem. Loss of organic matter primarily occurs through harvests and fires, but also after large pest and pathogen infestations, by animals browsing and grazing in intensive land use systems where animal faeces are not returned to the pastures. In all of these disturbances, biomass and associated nutrients are lost or exported from the ecosystem and/or decomposed somewhere else—for example, in human-dominated systems, in sewage works or by burning of waste. The heterotrophic organisms of the ecosystem in which the organic matter was once produced are no longer involved in the decomposition process.
Harvests and intensive grazing remove not only carbon as plant organic matter but also nutrients with the export of biomass. This export of nutrients can quickly lead to soil degradation if nutrients are not resupplied by fertilisation. Historically, litter raking in forests was an important forest use. Newly fallen leaves were collected and used during the winter as bedding for animals in stables and taken to the fields in spring as fertiliser. As leaves and needles have considerably higher nutrient contents than wood, this form of forest use drastically decreased forest soil fertility and led to reallocation of nutrients from forests to fields over large areas of land.
The effect of this type of forest use is still visible after decades as shallow humus layers in the affected forests (Schulze (2000), a site at Aubure in France). The impacts of intensive grazing strongly depend on the stocking rate of the livestock, which must be in accordance with the vegetative growth to be sustainable. If these sustainable stocking rates are exceeded, overgrazing occurs, which results first in a change of vegetation composition, then in a decrease of vegetation cover, then open (bare) soil and, finally, soil erosion—visible, for example, as tracks of grazing on steep slopes in alpine meadows. Measurement of net ecosystem CO2 fluxes over a pasture in Panama clearly demonstrated how overgrazing can lead to large CO2 losses, turning the pasture into a carbon source for the 2 years of the study (Wolf et al. 2011).
Another globally very important disturbance is fire, which can destroy standing biomass or deep peat soils over large areas in a short time. The intensity of the fire depends mainly on the available biomass but also on the wind speed, the moisture content of the fuel and oxygen concentrations (relevant for below-ground smouldering peat fires). Therefore, rare fires—in which plant biomass and soil organic matter that have accumulated in the long interval between the fires are consumed (stand-replacing fires)—can have more drastic effects than frequent and thus less intense ground fires. A famous example of such a fire is the one in Yellowstone National Park in 1988, which destroyed 320,000 ha over a couple of weeks because of special weather conditions and wildfire suppression for more than 50 years. Low- intensity fires (surface fires) are frequent in savanna systems, where temperatures at ground level usually remain below 100 °C and 40 °C is rarely reached at a 10 cm soil depth (Bradstock and Auld 1995). The often very short duration (only minutes) of the fire at the same site, owing to the low fuel load (i.e. biomass), is the reason for these fire characteristics.
Fire releases large amounts of carbon and nitrogen from the system in the form of different gaseous oxidation products (Konovalov et al. 2014), while base cations mostly remain in the ash in the ecosystem. A fraction of the remaining organically bound nitrogen is readily transformed into ammonium and nitrate in the warm soils covered with dark ash. Thus, seeds, young plants and surviving rhizomes experience good growth conditions, sometimes even better than those before the fire. Therefore, fire is used worldwide as an agricultural practice to burn off plant residues that would otherwise not decompose easily (e.g. in dry and hot areas) and to support new growth by increasing nutrient availability right after the fire. However, in the long-term, fire will lead to a reduction of soil fertility, particularly for the N supply, if this resource loss is not counteracted either by legumes or by fertilisation.
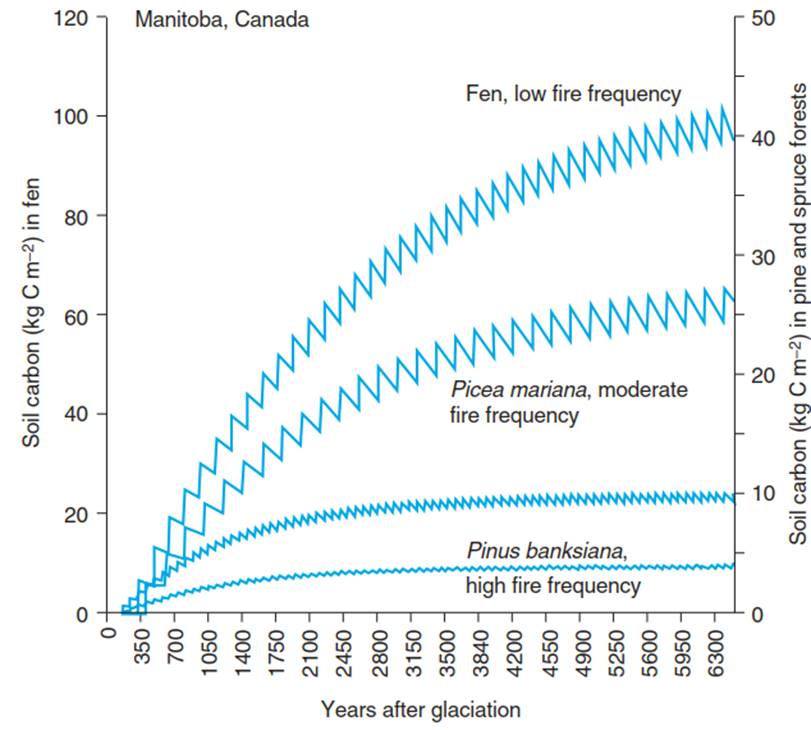
Fig. 13.7. Modelled changes in soil carbon in a wetland fen with a low frequency of fires and in Picea mariana and Pinus banksiana stands experiencing increased fire frequencies. The zig-zags of the curves reflect the frequency with which fires occurred. The figure illustrates the cumulative amount of C that is removed from these ecosystems directly or indirectly by fires (Harden et al. 2000)
In boreal coniferous forests, the fire sequence determines the amount of carbon that may be stored in the soil (Fig. 13.7). Calculated over a period of 6000 years (since the retreat of the Laurentide Ice Sheet in North America), the frequently burned Pinus banksiana stands contain only half as much C in the soil as the less frequently burned Picea mariana stands. These amounts, in turn, are significantly smaller than those in very rarely burned blanket bogs where, independently of the fire frequency, the high water table conserves the carbon in the soil, indicating this ecosystem’s potential to accumulate organic matter. Saturation occurs in all cases—that is, there is a balance between the formation of organic matter in the soil and its consumption by respiration and fires—but this balance is also affected by the high frequency of fires and the low productivity of the vegetation (Harden et al. 2000).
Date added: 2025-02-05; views: 449;