The Environment and Organism Are Tightly Coupled
Continuous, reciprocal involvement between organism and environment means that the environment does more than merely set the evolutionary stage for the organism. The environment and organism together establish the ‘fitness function, in other words, the success or failure of the phenotype within a selective milieu. The environment plays a role in developmental processes thereby influencing the phenotype itself (Fig. 7.1 and Scheiner 1993).
Lewontin (1983, 2000) has made the essence of the points shown in Fig. 7.1 clear in two forceful analogies as well as several corollaries (Table 7.1). First, the developmental paradigm that genetically predestined ‘normal’ ontogeny simply unfolds against a conducive environmental backdrop—much as photographic film would be converted from latent image to image in the appropriate developing solution—is wrong. Rather, the organism is both cause and consequence of a particular ontogenetic sequence. There is no overall best phenotype; what is best depends on the environmental context.
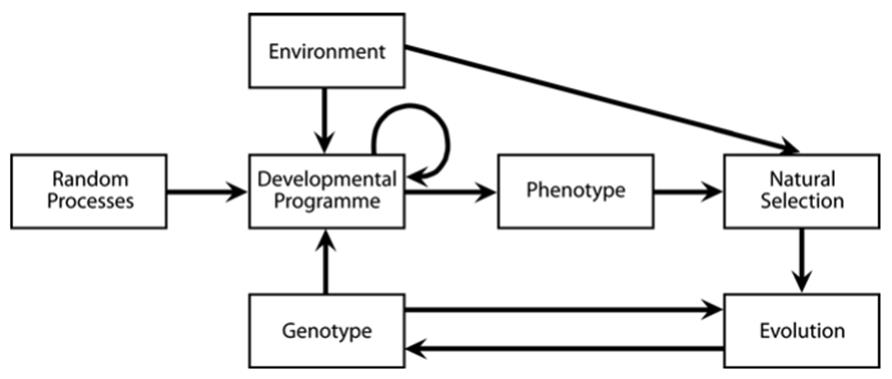
Fig. 7.1. Model illustrating how environment interacts with genotype (and random events) to influence the developmental program and, ultimately, to shape the variable or plastic phenotype. The environment also drives natural selection by determining in part the survivors and is, in turn, affected by the composition of the resulting population and community. Note also that genetic effects on phenotype can extend beyond the traditional phenotype level to become manifest at higher organizational levels (Whitham et al. 2006), producing what have become known as 'community and ecosystem phenotypes'. From Scheiner (1993). Reproduced from Annual Review of Ecology and Systematics by permission of Annual Reviews Inc. ©1993

Table 7.1. The interaction of organism with environment: Some postulates (Lewontin 1983, 2000)
This is dramatically shown in the classic experiment of Clausen et al. (1948) of differential performance of clonal transplants of Achillea grown at three elevations in California. The stature of each [genetically identical] clone varied more-or-less extensively as a function of elevation. Second, Lewontin argues that the phylogenetic paradigm wherein the environment poses successive ‘problems,’ which each species ‘solves’ adaptively, also is mistaken. Each species, indeed each organism, is a unique product of a genotypic and environmental history, not to mention random events (later). In Lewontin’s words (1983, p. 276), noted at the outset of this chapter, “...genes do not determine individuals nor do environments determine species.” By extrapolation this implies that evolution is the concerted change of both organism and environment.
There is probably no more elegant example of the organism creating its own environment than that of the tumor- inducing plant bacterium Agrobacterium tumefaciens. As part of the complex signaling process between the pathogen and host, and ensuing pathogenesis, the bacterium, which resides in the root, genetically engineers the plant to produce novel, specific metabolites (opalines) useful to itself (see discussion in Chap. 3 and McCullen and Binns 2006; Platt et al. 2014). In effect, it has obtained the keys to the pantry and has locked itself inside.
Recognizing that the association between organism and environment is dynamic, reciprocal, and continuous invigorates the longstanding dogma that environment and genetics together determine the individual. As an example, we might consider some interesting aspects of the life history of the remarkable coastal redwood tree (Sequoia sempervirens) in its many phenotypic representations (Figs. 7.2 and 7.3). This redwood species has been measured at heights commonly of 100 m or more and extreme longevity, the oldest confirmed tree being at least 2200 years (Noss 2000). Present-day stands of the species are confined by the environment to a narrow strip of the Pacific coastline extending some 700 km from southwestern Oregon to Monterey County south of San Francisco (Noss 2000). These favorable conditions result from a set of unique circumstances, principally high summer humidity and free moisture associated with frequent oceanic fogs, continuously cool temperatures, low evapotranspiration, and deep, moist, especially alluvial soils.
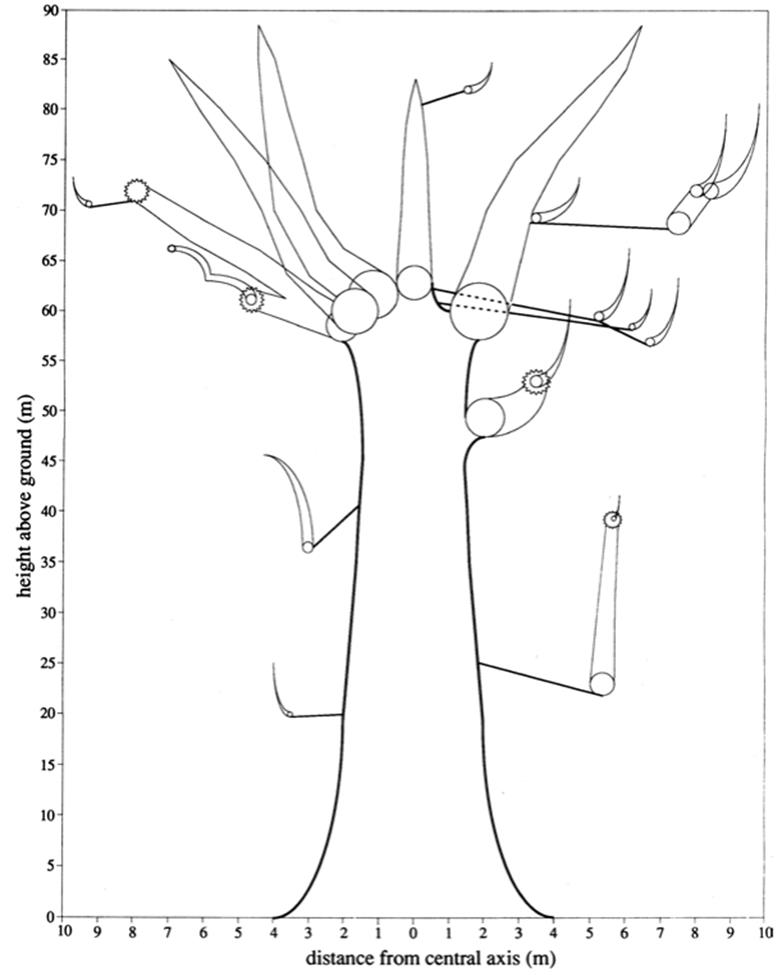
Fig. 7.2. Phenotypic model at a moment in time of a remarkable organism as shaped by the environment: The 'Atlas Tree', the fifteenth largest known living coastal redwood at 88.6 m high, 7.1 m diameter at breast height (dbh), and estimated wood volume of 791 m3, located in Prairie Creek Redwoods State Park, Humboldt County, California. Note expanded scale of x-axis. The irregular conical lines represent branches; only branches giving rise to 'reiterated' trunks (those arising after breakage of a main axis) are shown. There are 21 such trunks. Circles denote girth of trunks and branches and are drawn to same scale as the x-axis. Jagged-edged circles represent broken trunks. See Chap. 4 in Noss (2000). From The Redwood Forest (2000) edited by Reed F. Noss, copyright ©2000 by Save-the-Redwoods League. Reproduced by permission of Island Press, Washington, DC. Compare with Fig. 7.3

Fig. 7.3. A scale drawing of the 'Atlas Tree' shown in Fig. 7.2 (drawing by R. Van Pelt). Note the complexity of the crown featuring the multiple trunks. The specks at the base of the tree represent humans and a dog at the same scale as the tree. From The Redwood Forest (2000) edited by Reed F. Noss, Copyright ©2000 by Save- the-Redwoods League. Reproduced by permission of Island Press, Washington, DC
The atmospheric conditions in turn hinge largely on the broader climatic setting, which features a coastal upwelling system driven by wind (The California Current; see Black et al. 2014). While fog is rightly implicated universally as a key factor in the tree’s habitat, it is a necessary but insufficient component. Redwoods are virtually absent from coastal areas where soils are not supportive, such as within a sector immediately west of the San Andreas Fault line at Point Reyes, California, whereas the trees are common on an adjoining sector with a different soil type immediately east (Shuford and Timossi 1989). Interception of fog provides a substantial portion of the water budget of the tree (Dawson 1998), as well as input as fog drip to the diverse understory plant and animal communities intimately associated with both living and dead redwood biomass.
Fog relieves moisture stress characteristic of canopies at such heights related to challenges of water conduction from the roots to uppermost leaves (for the physiological and hydraulic implications, see Givnish et al. 2014; Koch et al. 2004, 2015; Klein et al. 2015). Nevertheless, the morphology and anatomy of leaves near the bottom of trees (large, expanded) are dramatically different from those near the top (small, scale-like), attributable mainly to reduced turgor (water potential) with height (Koch et al. 2004). These differences, in turn, affect many processes, including photosynthesis. In other words, the leaves have substantially different ecophysiology and effectively live in dramatically different environments.
Older coastal redwoods typically have a complicated canopy structure marked by ‘reiterations’ (multiple trunks arising when the main axis breaks in storms and sprouts new trunks). Multiple fusions also occur among branches and from trunk-to-trunk and branch-to-trunk from branch-to-branch; such fusions can themselves resprout (as do fallen branches or trunks on the ground) with each reiterated trunk supporting its own branch system. Remarkably, even non-fused and completely severed branches lodged in the crown can remain alive and resprout (see Figs. 7.2 and 7.3 and Noss 2000). This aerial scaffold in turn supports a complex community of epiphytes, including microorganisms, lichens, bryophytes, and vascular plants, as well as animals (Noss 2000; Williams and Sillett 2007). The base of the tree also provides a complex habitat for multiple organisms, as does dead wood on the forest floor. In short, clearly the coastal redwood is the epitome of an organism that both shapes and is itself dramatically shaped by its environment.
Finally, it is noteworthy that organisms can differ in phenotype for reasons beyond both conventional genetics and environment. This random component of variation is known as noise (Lewontin 2000; Griffiths et al. 2015). For example, the number of eye facets in Drosophila is typically different between the left and right eye. As the eyes of the same individual are genetically the same and their environments are effectively identical, such differences almost certainly result from stochasticity operating at the molecular level during ontogeny. Even clonal bacterial cells growing in homogeneous (shake-culture) conditions display nongenetic individuality attributable to fluctuation of key molecules in such low quantities that they are subject to statistical variation (Spudich and Koshland 1976; Avery 2006).
The phenomenon is typically referred to as “phenotypic heterogeneity” in bacteriology (Ackermann 2015). In fact, noise arises from stochastic events both extrinsic and intrinsic to a given cell (Elowitz et al. 2002). Although it has been recognized for decades, only relatively recently has the far-reaching impact of noise and stochasticity become more fully appreciated. This is attributable to expanding interest in epigenetics and the rapidly advancing technology to dissect genetic and metabolic circuits, often at the level of individual molecules in living cells, (e.g., Losick and Desplan 2008; Eldar and Elowitz 2010). We discuss several examples and implications in the ensuing sections. It should further be noted that, notwithstanding the vagaries and versatility imposed by noise and the environment, ontogeny can be strongly genetically driven and adaptive. For example, Holeski et al. (2009) show in a common garden experiment with ramets propagated from the top, middle, and bottom of cottonwood trees that resistance to aphids is significantly higher in the top than bottom, i.e., that it is genetically programmed and not a function of environmental factors that vary with height such as sunlight or water relations.
Date added: 2025-06-15; views: 379;