How Organisms Experience Environments
The Influence of Life Span and Growth Form.In biological terms, there are no absolutely constant environments either in nature or even in the laboratory. Differences occur in space from minute increments to vast reaches (<µm to >km); likewise, cyclic and noncyclic changes occur through time (milliseconds at one extreme to eons on a geological scale). For instance, with respect to space, soil properties vary over several orders of magnitude influencing microbial distribution and activity at one extreme, to plant distribution patterns over regional scales at the other extreme. A similarly vast spectrum is found with respect to micro- and macro-variation in nutrients within the ocean extending from the microhabitat surrounding particles of marine detritus (Alldredge and Cohen 1987; Poindexter 1981a, b) to the nutrient upwellings along coasts or impacts of the Gulf Stream.
Temporal fluctuations range from the recurrent seasonal and diurnal cycles, to the highly irregular and episodic. Conducive environments may be fleeting, as in a shifting sand bar in the Mississippi River on a scale of years or decades for plant colonists; or phases in host susceptibility that change over minutes, hours, or days for plant and some animal pathogens, or on a geological timescale if we are considering continental drift and ancient phylogenies or global warming/cooling cycles. Such vastly different terms of reference frame the important question: At what threshold does environmental fluctuation become significant to the organism and what life history attributes most determine the response?
Each creature experiences the environment differently. A major part of this difference relates to growth form and longevity, including their correlates. A shorter lifespan, generally speaking, means a greater acuity to short-term fluctuations. Variability occurring at intervals beyond the typical lifespan is beyond the frame of reference of the organism (although obviously not of the lineage). Clonal organisms, as noted previously (Chap. 5), persist and expand indefinitely. The clonal unit (ramet) may only experience a particular environment over a short period (possibly minutes or hours in the case of a single bacterial cell; see example below), whereas the clone as a whole may persist for decades or centuries. Hence, as a genetic individual, the clone typically experiences over time greater ranges in environmental variables than does the genet in most unitary organisms. Also, by virtue of their size and whether aggregated or disaggregated, clones frequently experience multiple environments simultaneously.
Among unitary organisms, for a rotifer with a life of 10 days, weekly changes are probably about comparable in periodicity to yearly changes for a bird (Chap. 1 in MacArthur and Connell 1966). The life cycles of animals in the Florida Everglades are coupled to the annual periodicity of the wet and dry seasons. The wood stork breeds when water levels are falling and it can easily obtain fish in the receding pools. The bird will not nest if this water cycle is disrupted (Kahl 1964). Analogously, in the annual cycle for many fungal pathogens of plants, the apple-scab fungus Venturia inaequalis overwinters as a saprophyte in apple leaf litter on the orchard floor where it undergoes sexual recombination; it then oversummers as a parasite, undergoing repeated rounds of asexual fragmentation in association with the living plant. Both phases are intimately tied to the seasonal activities of the host as well as the orchard environment (Andrews 1984, 1992; Aylor 1998). In terms of a shorter time scale, leaf-associated bacterial populations respond quantitatively and qualitatively on the order of minutes to hours as physical conditions change (Hirano and Upper 1989, 2000). Growth rates over brief periods in nature can even be on an order of magnitude (doubling times ca. 2-3 h) approaching those under optimal laboratory conditions.
There are various biological recorders of ambient conditions. Perhaps the best known of these is variation in the width of annual growth rings of trees, which reflects the impact of many environmental variables, particularly water. This living chronograph can extend over thousands of years in the case of extraordinarily long-lived species such as bristle- cone pine (Pinus longaeva), among others (Pilcher et al. 1984). Thus, long-term oscillations in climate also are captured. A remarkable example is that growth rings of the blue oak (Quercus douglasii) projected back to the year 1428 have been used to infer climate pattern in the southwestern U.S. (Black et al. 2014). Such exhaustive reconstruction is possible because the same atmospheric conditions that control winter-time coastal upwelling (and associated marine life) in the California Current, noted earlier, also affect precipitation and tree growth.
For species comparison purposes, one can array various life spans or generation times against a set of hypothetically important environmental temporal variables (Fig. 7.4 and Istock 1984). Every organism experiences many variables concurrently, and differences in life history mean differences in aggregate experience with the environment. Organisms such as annual plants with a short life span relative, say, to a yearly pattern of seasonal change, are restricted to one season and become specialized to it such that they could not develop at other seasons. Species that live over many seasons could be generalists—doing more or less well throughout the year, or specialists—flourishing in one season while remaining dormant or migrating in others (see Dormancy below).
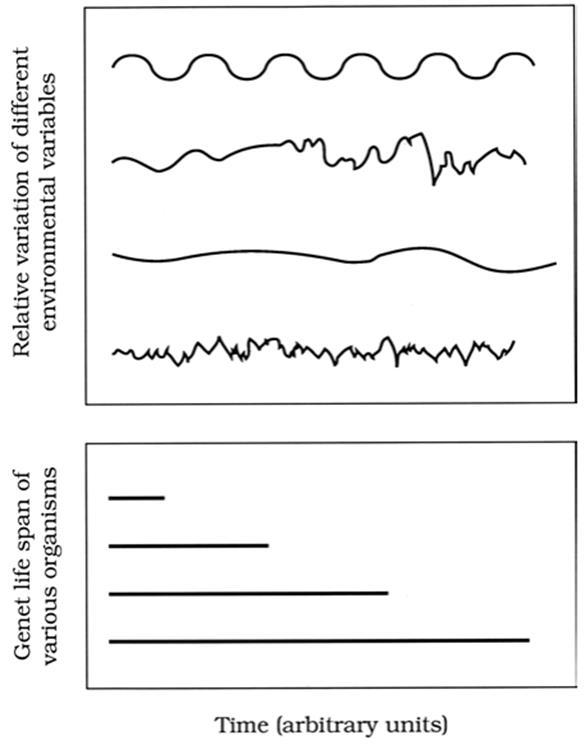
Fig. 7.4. Hypothetical periodicities of various abiotic environmental variables (top) arrayed against life spans or generation times of organisms. Variation also occurs in three-dimensional space as well as in time (not shown)
Date added: 2025-06-15; views: 384;