Cope's rule and the evolution of larger organisms
There has been an increase in size during evolution in the sense that among biota as a whole both the mean and upper size limits have increased through geological time (Carroll 2001; for synopsis see Chap. 2 in Bonner 1988). For example, Payne et al. (2009) compared the largest known fossil organisms from the origin of life in the Archaean (4000-2500 Mya) through the Phanerozoic (542-0 Mya) eons and found that maximum size as represented by their data set (cf. McShea 2001) increased 16 orders of magnitude over that approximate 3.5 billion years (Fig. 4.7). Most of this increase in size occurred in two episodes; not surprisingly, the first was associated with the advent of the eukaryotic cell and the second with multicellularity.
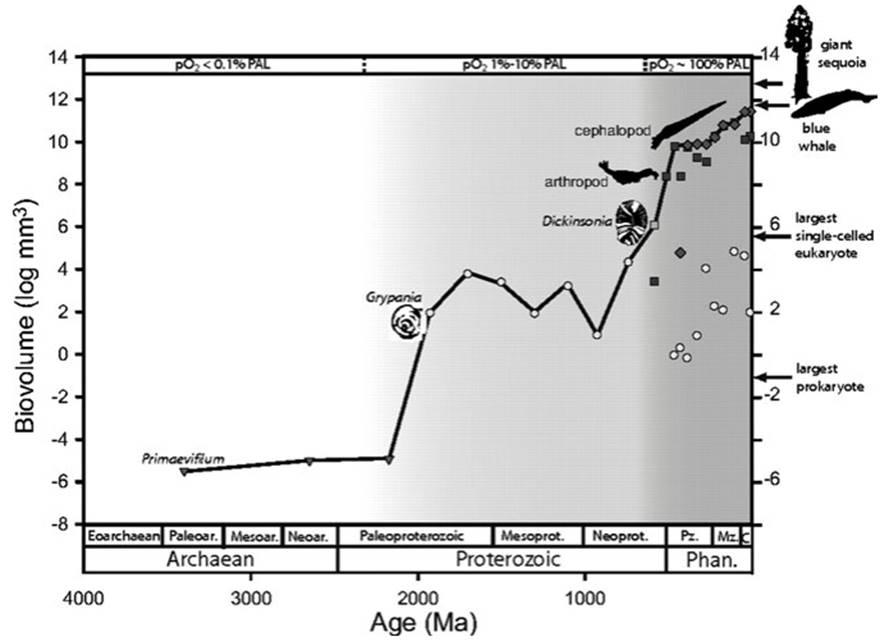
Fig. 4.7. Size plot of the largest fossils over geological time, with representative examples of unicellular eukaryotes, animals, and vascular plants. Sharp increases in size correspond approximately to increases in oxygen levels (note top axis). Symbols represent: triangles prokaryotes; circles protists; squares animals, except for the square labeled Dickinsonia, which represents pre-Cambrian, enigmatic, multicellular eukaryotes called vendobionts; diamonds vascular plants. On top axis, PAL = present-day values. Note authors' caveats about organisms included and what is meant by size of the 'individual' [size here excludes colonial organisms; for unicellular organisms, it is cell size, not clone size]. From Payne et al. (2009); reproduced from Proceedings of the National Academy of Sciences by permission of the National Academy of Sciences, USA
The above generality does not mean that the paleontological record is without periods or lineages of size decrease, and the rate of change of size tends to vary inversely with the duration over which it is studied (Gingerich 1983). For instance, organisms such as the birds, amphibians, and rotifers are smaller than their ancestors, and at least some of the extant unicellular fungi apparently are derived secondarily from multicellular ancestors.
The case for birds is particularly interesting. Extinction was not the fate of all the dinosaurs because extant birds share a common ancestor with them. This ancestral creature appears to have been perhaps 1-m or more long, some 20-30 kg in mass, and a bipedal carnivore of the early Triassic Period (ca. 250 Mya). While the other dinosaur clades were marked by size increase and in some cases to titanic proportions, the only sustained trend to size decrease was in the evolution of the avian lineage (Sereno 1999). Likewise, the modern club mosses (lycopods) are a few centimeters high compared with their ancestors, many meters in height, which proliferated in the luxuriant Carboniferous coal forests of 300 million years ago.
The tendency for most animal lineages to evolve towards large body size is epitomized by Cope’s Rule, in honor of the paleontologist Edward Drinker Cope who did not enunciate it as such but the principle emerged from his work during the late 1800’s (Stanley 1973; McKinney 1990). An important caveat is that Cope’s studies were focused on mammals over the Cenozoic (65 Mya-present) and he ascribed the mechanism specifically to an active, directional replacement of older (smaller) forms by larger ones under the influence of natural selection. Thus, the minimum size also increases and the explanation implies that larger organisms would be fitter. Lineages can also increase in body size by a passive trend, however, without unidirectional selection (see later comments on Gould’s [1996] argument for random “diffusion off the left wall”). In this case, larger descendants arise but the lower size category characteristic of the ancestors still remains, i.e., trait variance increases (Gould 1996; Carroll 2001).
So, while lineages generally increase in size and complexity through geological time, this is not invariable, and there is considerable debate over whether an active or passive mechanism is primarily operative in a given instance and whether there is also a wall on the right (see Knoll and Bambach 2000). Jablonski (1996) argued that for species of various molluscs in the late Cretaceous that there was an active trend for both size increase and size decrease. In a major critique of Cope’s Rule, Stanley (1973) began with the observation that if net size increase has prevailed in evolution, a prediction, which is borne out as a generality, would be that most higher taxa should have arisen at small (relatively unspecialized) body size relative to the optimum for the group. Larger (more specialized) taxa subsequently emerge, typically during speciation events, and the descendants fill new adaptive zones.
Thus, a size increase or size decrease actually may occur, depending on whether the mean size of the original species is smaller or larger than the optimum for the occupied niche. Major adaptive breakthroughs would tend to be restricted to small body sizes because large species typically are locked into a morphologically specialized body plan dictated by space:volume scaling constraints (following section), which is inherently unsuited for descendent taxa specialized in different ways (Stanley 1973).
Date added: 2025-06-15; views: 356;