Diffusion Across Biological Membranes: Mechanisms, Ion Channels, and Electrochemical Gradients
The distance a molecule must diffuse is a critical determinant of its transport rate, whether traveling from blood to a cell or moving within a cell's interior. Despite high individual molecular speeds, incessant collisions prevent straight-line travel, causing diffusion times to increase with the square of the distance. Consequently, while diffusion equilibrium is achieved rapidly over microscopic, cellular distances, it becomes impractically slow over macroscopic scales of just a few centimeters.
For large organisms like humans, relying solely on diffusion to transport oxygen and nutrients from the body surface to internal tissues would be fatally inadequate. This limitation is overcome by the circulatory system, which utilizes bulk flow—a pressure-driven mechanism (powered by the heart) for long-distance rapid transport. Diffusion remains essential for the short-distance movement of substances between the blood, interstitial fluid, and intracellular fluid.
Diffusion Through Membranes. While the principles of diffusion apply to solutes in water, biological transport often occurs across cellular membranes. Substances like metabolic wastes exiting cells or nutrients entering must traverse the plasma membrane. The membrane's presence significantly modulates the diffusion rate, measured by how quickly intracellular concentration equilibrates with the extracellular fluid.
Assuming a constant extracellular concentration due to its large volume (Figure 4.4), the net flux (J) across the membrane proceeds from high to low concentration. Its magnitude is governed by a modified form of Fick’s first law of diffusion:
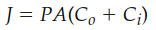
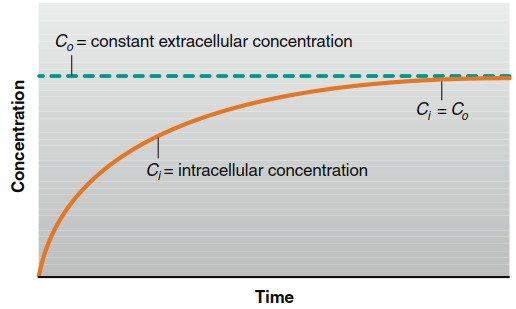
Figure 4.4. The increase in intracellular concentration as a solute diffuses from a constant extracellular concentration until diffusion equilibrium (Ci = Co) is reached across the plasma membrane of a cell.
Here, P is the membrane permeability coefficient, A is the surface area, and (C₀ - Cᵢ) is the concentration gradient across the membrane. The permeability coefficient is an experimental measure of a specific molecule's ease of passage through a given membrane at a set temperature. Molecules typically diffuse thousands to millions of times slower through membranes than through an equivalent thickness of water, establishing membranes as selective barriers.
Diffusion Through the Lipid Bilayer. Permeability correlates strongly with molecular structure. Nonpolar molecules (e.g., oxygen, carbon dioxide, steroid hormones) diffuse rapidly through the membrane's lipid bilayer because they readily dissolve in its hydrophobic, fatty acid interior. Conversely, polar molecules and ions have very low lipid solubility and thus cross slowly, if at all. This differential permeability illustrates a core principle: physiological processes are dictated by the laws of chemistry and physics. Many metabolic intermediates are polar or ionized and are thus retained within cellular compartments unless specialized transport proteins are present.
Diffusion of Ions Through Ion Channels. Ions like Na⁺, K⁺, Cl⁻, and Ca²⁺ cross membranes far faster than their minimal lipid solubility would predict, and permeabilities vary greatly between cell types. This indicates a protein-mediated pathway. Integral membrane proteins form aqueous pores called ion channels (Figure 4.5). These channels have narrow diameters, slightly larger than the ions themselves, ensuring selectivity.
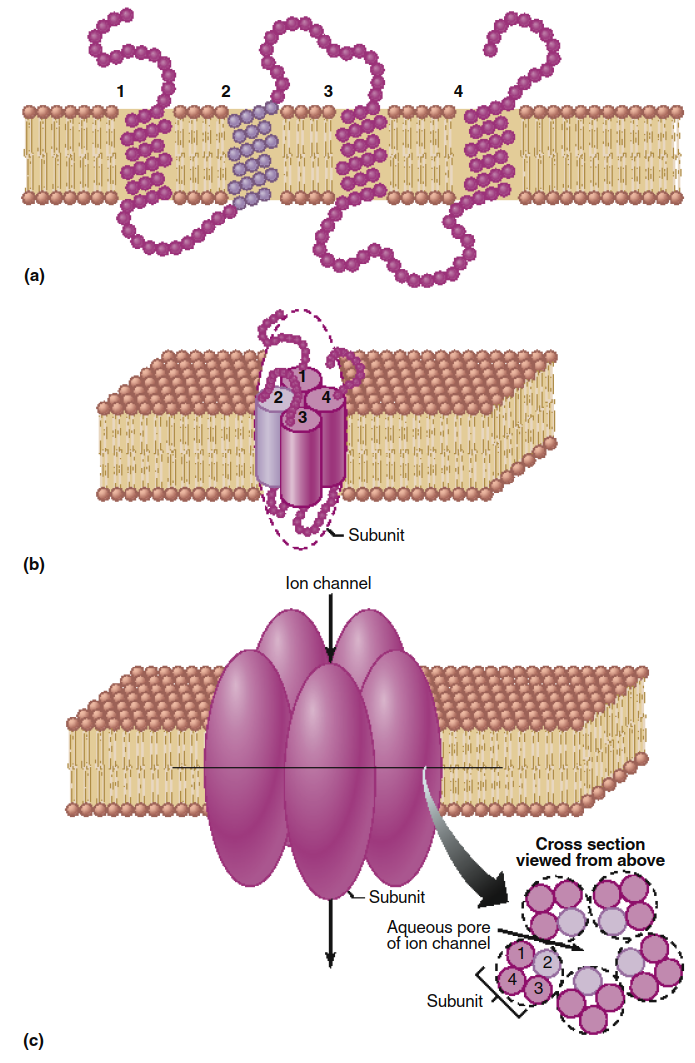
Figure 4.5. Model of an ion channel composed of five polypeptide subunits. Individual amino acids are represented as beads. (a) A channel subunit consisting of an integral membrane protein containing four transmembrane segments (1, 2, 3, and 4), each of which has an alpha-helical configuration within the membrane. Although this model has only four transmembrane segments, some channel proteins have as many as 12. (b) The same subunit as in (a) shown in three dimensions within the membrane, with the four transmembrane helices aggregated together and shown as cylinders. (c) The ion channel consists of five of the subunits illustrated in (b), which form the sides of the channel. As shown in cross section, the helical transmembrane segment 2 (light purple) of each subunit forms each side of the channel opening. The presence of ionized amino acid side chains along this region determines the selectivity of the channel to ions. Although this model shows the five subunits as identical, many ion channels are formed from the aggregation of several different types of subunit polypeptides.
Ion channels exhibit selectivity for specific ions based on pore size, the charge distribution on their lining, and interactions with an ion's waters of hydration. For instance, K⁺ channels are selectively permeable to potassium, while Na⁺ channels are selective for sodium. Thus, two membranes with equal numbers of K⁺ channels have equal potassium permeability but may differ vastly in sodium permeability based on their Na⁺ channel content.
Effects of Electrical Forces on Ion Movement. For ions, the driving force for diffusion includes both chemical (concentration) and electrical forces. A membrane potential—a voltage difference across the plasma membrane (Figure 4.6)—exerts an electrical force on charged particles. This creates an electrochemical gradient, the combined influence of the concentration gradient and the membrane potential.
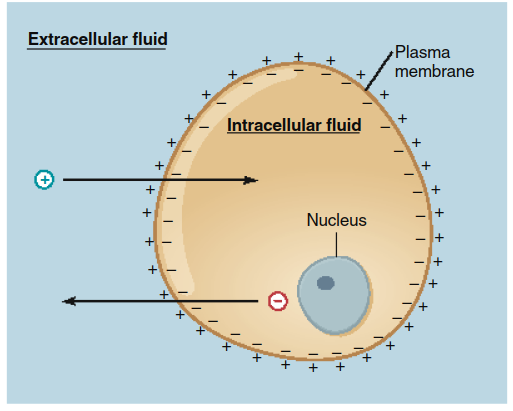
Figure 4.6. The separation of electrical charge across a plasma membrane (the membrane potential) provides the electrical force that tends to drive positive ions (1) into a cell and negative ions (2) out.
These forces can act in concert or opposition. For example, a membrane potential might attract K⁺ into the cell while its concentration gradient drives it out. The net ion flux is determined by the net electrochemical gradient.
Regulation of Diffusion Through Ion Channels. Ion channels are dynamic, transitioning between open and closed states (Figure 4.7) in a process called channel gating. The total ion flow depends on how often and how long a channel opens. Channel conformation can be regulated by three primary mechanisms:
1. Ligand-Gated Ion Channels: The binding of a specific ligand (e.g., a neurotransmitter) induces an allosteric or covalent change in the channel protein, opening or closing it.
2. Voltage-Gated Ion Channels: Changes in the membrane potential cause charged regions of the channel protein to move, altering its shape.
3. Mechanically Gated Ion Channels: Physical deformation or stretching of the membrane changes the channel's conformation.
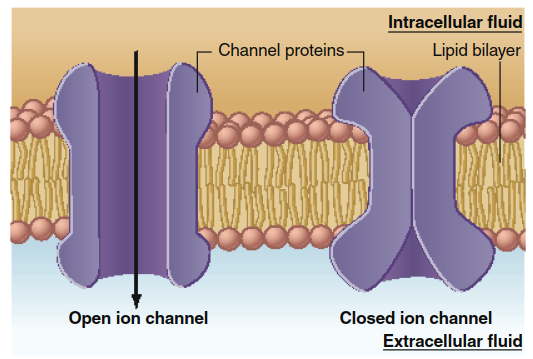
Figure 4.7. As a result of conformational changes in the proteins forming an ion channel, the channel may be open, allowing ions to diffuse across the membrane, or may be closed. The conformational change is grossly exaggerated for illustrative purposes. The actual conformational change is more likely to be just sufficient to allow or prevent an ion to fit through.
A single ion type, such as K⁺, may traverse via multiple channel types (ligand-gated, voltage-gated, mechanically gated), each playing distinct roles in cell signaling, communication, and electrical excitability, as explored in later chapters. This gating capability allows cells to precisely and rapidly control their permeability and electrochemical state.
Date added: 2026-07-14; views: 3;