Aquaporins and Osmosis: Water Transport Across Cell Membranes
Water, despite being a polar molecule, diffuses rapidly across the plasma membranes of most cells. This rapid diffusion is mediated by a family of membrane proteins known as aquaporins, which form specialized channels through which water molecules can pass. The type and number of these water channels vary significantly among different cellular membranes, resulting in differential water permeability across cell types. Moreover, in certain cells, the number of aquaporin channels—and consequently the membrane's permeability to water—can be dynamically altered in response to various physiological signals.
This regulatory mechanism is particularly important in the epithelial cells lining specific ducts within the kidneys. As will be explored in Chapter 14, one of the kidney's primary functions is to regulate the volume of water excreted in urine, thereby maintaining homeostasis of total body water within fluid compartments. The epithelial cells of kidney ducts contain numerous aquaporins, whose abundance can be increased or decreased based on the body's current water balance. For instance, during dehydration, the number of aquaporins in the membranes of kidney epithelial cells increases, facilitating additional water movement from the forming urine back into the bloodstream. This physiological response explains why urine volume decreases significantly when an individual becomes dehydrated.
Osmosis and Water Concentration Gradients. The net diffusion of water across a semipermeable membrane is defined as osmosis. Like all diffusion processes, osmosis requires a concentration difference to generate a net flux of water molecules. Understanding how a difference in water concentration can be established across a membrane requires examining the relationship between solutes and water molecules in solution.
The addition of a solute to pure water effectively decreases the concentration of water molecules within that solution. For example, when glucose is dissolved in water, the resulting solution contains fewer water molecules per unit volume compared to an equal volume of pure water, as each glucose molecule occupies space previously filled by a water molecule (Figure 4.16). In quantitative terms, one liter of pure water weighs approximately 1000 grams, and the molecular weight of water is 18 g/mol, yielding a water concentration of 1000/18 = 55.5 M in pure water. The decrease in water concentration within a solution is approximately equal to the concentration of added solute, meaning one solute molecule displaces approximately one water molecule.
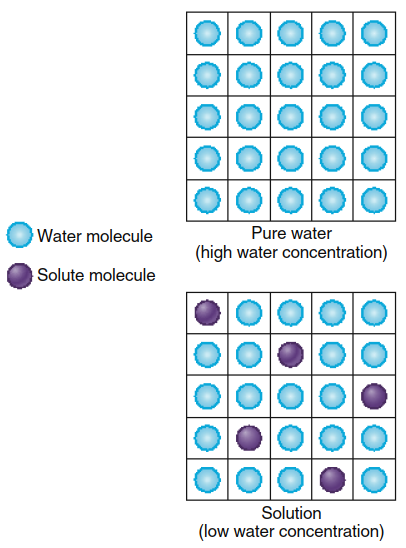
Figure 4.16. The addition of solute molecules to pure water lowers the water concentration in the solution
Therefore, the water concentration in a 1 M glucose solution is approximately 54.5 M rather than 55.5 M. Just as adding water dilutes a solute, adding solute effectively "dilutes" the water molecules in solution. This principle establishes that higher solute concentrations directly correlate with lower water concentrations within any given solution.
Solute Particles and Water Concentration. The degree to which water concentration decreases upon solute addition depends exclusively on the number of solute particles (molecules or ions) in solution, not on the chemical identity of the solute itself. For instance, 1 mole of glucose dissolved in 1 liter of solution decreases water concentration to the same extent as 1 mole of an amino acid, 1 mole of urea, or 1 mole of any other molecule that remains as a single particle in solution. However, molecules that ionize in solution decrease water concentration in proportion to the total number of ions generated upon dissociation.
Many simple salts dissociate nearly completely in aqueous solutions. For simplification, we assume 100% dissociation at body temperature and at concentrations typically found in blood. Consequently, 1 mole of sodium chloride (NaCl) in solution yields 1 mole of sodium ions (Na⁺) and 1 mole of chloride ions (Cl⁻), producing 2 moles of total solute particles. This reduces water concentration twice as much as 1 mole of non-ionizing glucose. Similarly, a 1 M magnesium chloride (MgCl₂) solution, if completely dissociated, would decrease water concentration three times more than a 1 M glucose solution, producing Mg²⁺ and two Cl⁻ ions.
Osmolarity: Total Solute Concentration. Because water concentration in a solution depends on the total number of solute particles, a concentration term that refers to the combined concentration of all solute particles—regardless of their chemical composition—proves highly useful. This total solute concentration is known as osmolarity. One osmol is defined as 1 mole of solute particles. Therefore, a 1 M solution of glucose has a concentration of 1 Osm (1 osmol per liter), whereas a 1 M solution of NaCl contains 2 Osm of solute per liter of solution.
A liter of solution containing 1 mole of glucose and 1 mole of NaCl has an osmolarity of 3 Osm. A solution with an osmolarity of 3 Osm could contain 1 mole of glucose plus 1 mole of NaCl, or 3 moles of glucose, or 1.5 moles of NaCl, or any other combination of solutes, provided the total solute concentration equals 3 Osm. Although osmolarity specifically quantifies solute particle concentration, it simultaneously determines the water concentration in the solution because higher osmolarity invariably means lower water concentration. Importantly, any two solutions sharing the same osmolarity will have identical water concentrations, as the total number of solute particles per unit volume is equal.
Osmosis Across Membranes. Applying these principles to osmosis across membranes, Figure 4.17 illustrates two 1-liter compartments separated by a membrane permeable to both solute and water. Initially, compartment 1 contains a solute concentration of 2 Osm, while compartment 2 contains 4 Osm. This solute concentration difference establishes a corresponding water concentration gradient: approximately 53.5 M water in compartment 1 and 51.5 M water in compartment 2. Consequently, a net diffusion of water occurs from the higher concentration in compartment 1 to the lower concentration in compartment 2, while solute diffuses in the opposite direction from compartment 2 to compartment 1.
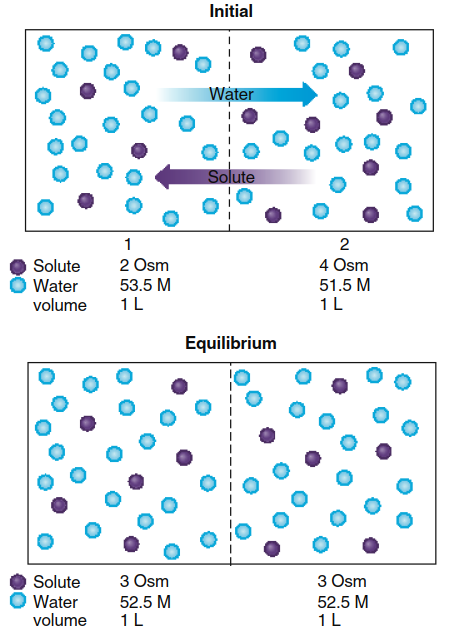
Figure 4.17. Between two compartments of equal volume, the net diffusion of water and solute across a membrane permeable to both leads to diffusion equilibrium of both, with no change in the volume of either compartment. (For clarity, not all water molecules are shown here or in Figure 4.18.)
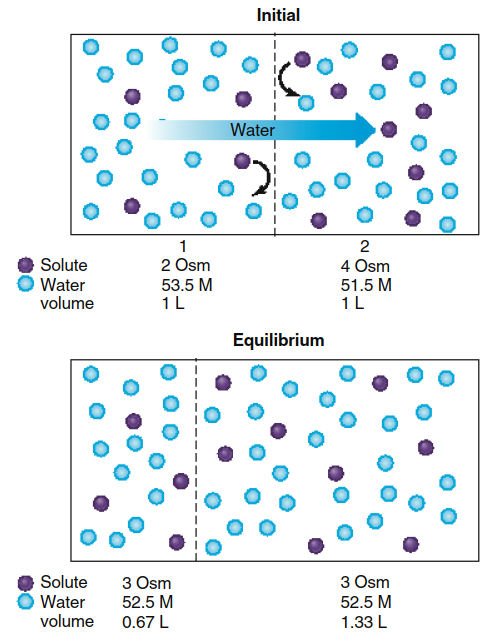
Figure 4.18. The movement of water across a membrane that is permeable to water but not to solute leads to an equilibrium state involving a change in the volumes of the two compartments. In this case, a net diffusion of water (0.33 L) occurs from compartment 1 to 2. (We will assume that the membrane in this example stretches as the volume of compartment 2 increases so that no significant change in compartment pressure occurs.)
At diffusion equilibrium, both compartments achieve identical solute and water concentrations of 3 Osm and 52.5 M, respectively. One mole of water diffuses from compartment 1 to compartment 2, while 1 mole of solute diffuses from compartment 2 to compartment 1. Because 1 mole of solute replaces 1 mole of water in compartment 1, and vice versa in compartment 2, no net volume change occurs in either compartment when the membrane is permeable to both water and solute.
When the membrane is replaced with one permeable to water but impermeable to solute (Figure 4.18), the same equilibrium concentrations of water and solute are eventually reached, but significant volume changes occur in both compartments. Water diffuses from compartment 1 to compartment 2, but no solute diffusion occurs in the opposite direction because the membrane blocks solute passage. Water continues diffusing into compartment 2 until water concentrations equalize on both sides of the membrane.
As water enters compartment 2, the solute concentration there decreases due to dilution, while solute in compartment 1 becomes more concentrated as water exits. When water reaches diffusion equilibrium, the osmolarities of both compartments become equal, meaning solute concentrations must also be equal. To achieve this equilibrium state, sufficient water must transfer from compartment 1 to compartment 2 to increase compartment 2's volume by one-third while decreasing compartment 1's volume by an equal amount. Critically, it is the presence of a membrane impermeable to solute that generates the volume changes characteristic of osmosis.
Osmotic Pressure. The two compartments in our example were treated as infinitely expandable, allowing net water transfer without creating a pressure difference across the membrane. However, if the walls of compartment 2 in Figure 4.18 had limited expansion capacity—as occurs across plasma membranes—water movement into compartment 2 would increase pressure within compartment 2, which would oppose further net water entry. Thus, water movement into compartment 2 can be prevented by applying pressure to compartment 2.
This leads to an important definition: when a solution containing solutes is separated from pure water by a semipermeable membrane (permeable to water but not to solutes), the pressure that must be applied to the solution to prevent net water flow into it is known as the osmotic pressure of that solution. The greater the osmolarity of a solution, the greater its osmotic pressure. It is crucial to recognize that osmotic pressure does not actively push water molecules into a solution; rather, it represents the amount of pressure required to prevent net water flow into the solution. Like osmolarity, osmotic pressure serves as a measure of a solution's water concentration—the lower the water concentration, the higher the osmotic pressure.
Extracellular Osmolarity and Cell Volume. The principles of osmosis can now be applied directly to cells, which satisfy all criteria necessary for producing osmotic water flow across a membrane. Both intracellular fluid and extracellular fluid contain water, and cells are enclosed by a plasma membrane that is highly permeable to water but impermeable to many substances. Substances unable to cross the plasma membrane are termed nonpenetrating solutes, meaning they do not diffuse through the lipid bilayer.
Most extracellular solute particles are sodium ions (Na⁺) and chloride ions (Cl⁻). Although these ions can diffuse into cells through ion channels or enter during secondary active transport, the plasma membrane contains Na⁺/K⁺-ATPase pumps that actively transport Na⁺ out of the cell. Therefore, while Na⁺ moves into cells, it is continuously pumped back out, effectively behaving as if it never entered. For this reason, extracellular Na⁺ functions as a nonpenetrating solute. Any chloride ions entering cells are similarly removed rapidly due to electrical repulsion generated by the membrane potential and the action of various transporters. Like Na⁺, extracellular chloride ions therefore behave as nonpenetrating solutes.
Inside the cell, the major solute particles are potassium ions (K⁺) and various organic solutes. Most organic solutes are large polar molecules unable to diffuse through the plasma membrane. Although K⁺ can diffuse out of cells through K⁺ channels, it is actively transported back inside by the Na⁺/K⁺-ATPase pump. The net effect, as with extracellular Na⁺ and Cl⁻, is that intracellular K⁺ behaves as a nonpenetrating solute confined to the intracellular fluid. Thus, Na⁺ and Cl⁻ outside the cell, along with K⁺ and organic solutes inside the cell, function as nonpenetrating solutes on opposite sides of the plasma membrane.
Tonicity and Cell Volume Regulation. The normal osmolarity of extracellular fluid ranges from 285-300 mOsm (rounded to 300 mOsm throughout this text unless otherwise noted). Because water freely diffuses across plasma membranes, water in the intracellular and extracellular fluids reaches diffusion equilibrium, resulting in equal osmolarities of approximately 300 mOsm in both compartments. Changes in extracellular osmolarity can cause cells—such as the red blood cells shown in the chapter-opening photo—to shrink or swell as water molecules move across the plasma membrane.
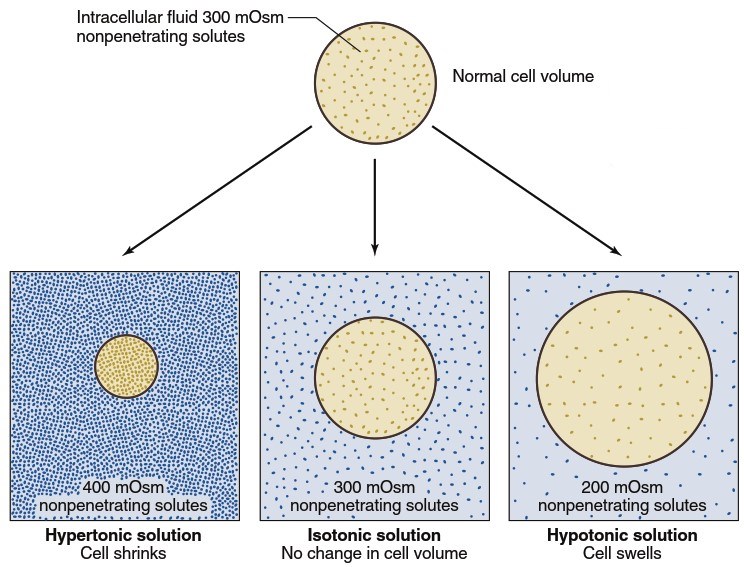
Figure 4.19. Changes in cell volume produced by hypertonic, isotonic, and hypotonic solutions
When cells with an intracellular osmolarity of 300 mOsm are placed in a solution of nonpenetrating solutes also having an osmolarity of 300 mOsm, they neither swell nor shrink because water concentrations are equal on both sides of the membrane, and solutes cannot enter or leave the cells. Such solutions are termed isotonic (Figure 4.19), meaning any solution that does not alter cell size. Isotonic solutions have the same concentration of nonpenetrating solutes as normal extracellular fluid. In contrast, hypotonic solutions have a nonpenetrating solute concentration lower than that found inside cells, causing water to move by osmosis into the cells, leading to cell swelling. Similarly, solutions containing greater than 300 mOsm of nonpenetrating solutes (hypertonic solutions) cause cells to shrink as water diffuses out of cells into the fluid with lower water concentration.
The concentration of nonpenetrating solutes in a solution—not the total osmolarity—determines its tonicity (isotonic, hypotonic, or hypertonic). By contrast, penetrating solutes that readily diffuse through lipid bilayers do not contribute to a solution's tonicity, as their concentrations rapidly equilibrate across the membrane.
Distinguishing Osmolarity from Tonicity. Another set of terms—isoosmotic, hypoosmotic, and hyperosmotic—denotes the osmolarity of a solution relative to normal extracellular fluid without regard to whether solutes are penetrating or nonpenetrating. These two sets of terms are therefore not synonymous. For example, a 1-liter solution containing 150 mOsm each of nonpenetrating Na⁺ and Cl⁻ plus 100 mOsm of urea (which rapidly crosses plasma membranes) would have a total osmolarity of 400 mOsm and would be hyperosmotic relative to a typical cell. However, this same solution would also be isotonic, producing no net change in the equilibrium volume of cells immersed in it.

Initially, cells placed in this solution would shrink as water moved into the extracellular fluid. However, urea, as a penetrating solute, would quickly diffuse into the cells until reaching the same concentration as in the extracellular solution. Consequently, both intracellular and extracellular solutions would soon achieve equal osmolarities, eliminating any difference in water concentration across the membrane and resulting in no net change in final cell volume. This occurs even though the extracellular fluid remains hyperosmotic relative to the normal value of 300 mOsm. Table 4.3 provides a comprehensive comparison of the various terms used to describe solution osmolarity and tonicity.
Date added: 2026-07-14; views: 4;