Endocytosis and Exocytosis: Cellular Transport Mechanisms Beyond Diffusion
In addition to diffusion and mediated transport, cells employ alternative pathways for substance transport that do not require molecules to traverse the structural matrix of the plasma membrane. When examined under an electron microscope, researchers frequently observe regions of the plasma membrane folding inward to form small pockets that subsequently pinch off, creating intracellular, membrane-bound vesicles containing a small volume of extracellular fluid. This fundamental process is termed endocytosis (Figure 4.20). The reverse mechanism, exocytosis, occurs when cytoplasmic membrane-bound vesicles fuse with the plasma membrane, releasing their contents into the extracellular space (see Figure 4.20).
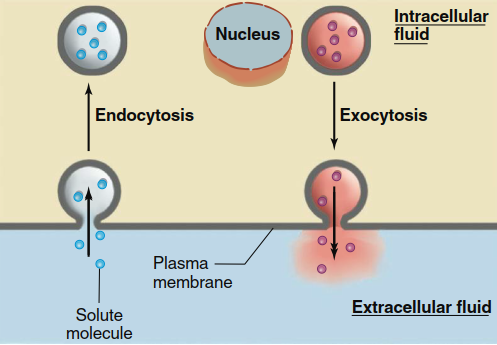
Figure 4.20. Endocytosis and exocytosis
Endocytosis. Three common types of endocytosis occur in eukaryotic cells: pinocytosis ("cell drinking"), phagocytosis ("cell eating"), and receptor-mediated endocytosis (Figure 4.21). Each mechanism serves distinct physiological functions and involves different molecular machinery. Understanding these processes is essential for comprehending how cells acquire nutrients, defend against pathogens, and maintain membrane homeostasis.
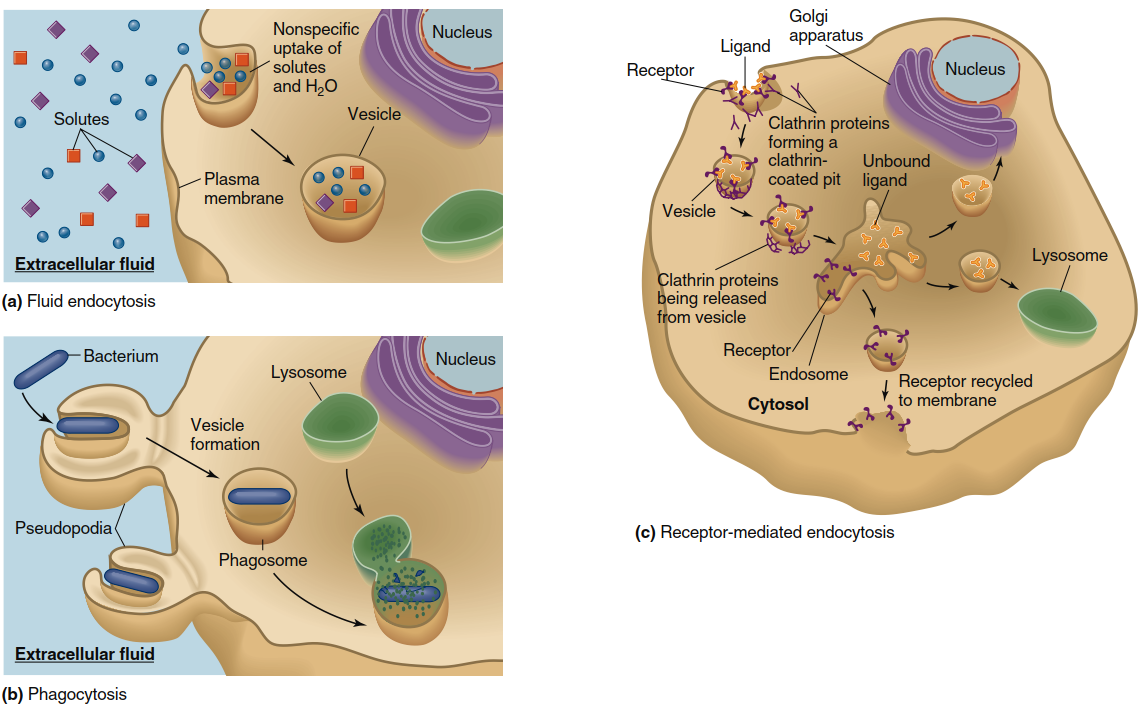
Figure 4.21. Pinocytosis, phagocytosis, and receptor-mediated endocytosis. (a) In pinocytosis, solutes and water are nonspecifically brought into the cell from the extracellular fluid via endocytotic vesicles. (b) In phagocytosis, specialized cells form extensions of the plasma membrane called pseudopodia, which engulf bacteria or other large objects such as cell debris. The vesicles that form fuse with lysosomes, which contain enzymes and other molecules that destroy the vesicle contents. (c) In receptor-mediated endocytosis, a cell recognizes a specific extracellular ligand that binds to a plasma membrane receptor. The binding triggers endocytosis. In the example shown here, the ligand-receptor complexes are internalized via clathrin-coated vesicles, which merge with endosomes (for simplicity, adapter proteins are not shown). Ligands may be routed to the Golgi apparatus for further processing, or to lysosomes. The receptors are typically recycled to the plasma membrane
Pinocytosis. In pinocytosis, also referred to as fluid endocytosis, an endocytotic vesicle encloses a small volume of extracellular fluid in a nonspecific manner. This process lacks molecular selectivity because the vesicle simply engulfs the aqueous extracellular fluid along with any dissolved solutes present, including ions, nutrients, and other small extracellular molecules. Large macromolecules, entire cells, and cellular debris do not typically enter cells via pinocytosis, making this pathway suitable primarily for sampling the extracellular environment and acquiring dissolved nutrients.
Phagocytosis. During phagocytosis, cells engulf bacteria or large particles such as cellular debris originating from damaged tissues. In this endocytotic variant, plasma membrane extensions called pseudopodia fold around the target particle's surface, completely surrounding it before internalization. The pseudopodia, containing their engulfed contents, subsequently fuse to form large vesicles known as phagosomes, which become internalized within the cell. These phagosomes migrate toward and fuse with lysosomes in the cytoplasm, where the phagosomal contents are destroyed by lysosomal enzymes and other degradative molecules. While most cells can perform pinocytosis, only specialized cell types—particularly those belonging to the immune system (Chapter 18)—carry out phagocytosis as a primary defense mechanism against pathogens.
Receptor-Mediated Endocytosis. In contrast to the relatively nonselective processes of pinocytosis and phagocytosis, most cells possess the capacity for specific uptake of molecules essential for cellular function or structural integrity. In receptor-mediated endocytosis, specific molecules present in the extracellular fluid bind to particular proteins located on the plasma membrane's outer surface. These specialized proteins are called receptors, and each receptor recognizes one ligand with high binding affinity (see Section C of Chapter 3 for detailed discussion of ligand-protein interactions). In one well-characterized form of receptor-mediated endocytosis, the receptor undergoes a conformational change upon ligand binding, initiating a cascade of molecular events.
Through sequential steps, a cytosolic protein called clathrin is recruited to the plasma membrane, where a class of adaptor proteins links the ligand-receptor complex to clathrin. The entire complex then assembles into a cage-like structure that promotes aggregation of ligand-bound receptors into a localized membrane region, forming a depression known as a clathrin-coated pit. This pit subsequently invaginates and pinches off to generate a clathrin-coated vesicle. By localizing ligand-receptor complexes to discrete membrane patches before endocytosis, cells obtain concentrated amounts of specific ligands without engulfing large volumes of extracellular fluid from multiple membrane sites. Consequently, receptor-mediated endocytosis achieves selective concentration of particular ligands within the endocytotic vesicle.
Intracellular Fate of Endocytotic Vesicles. Once an endocytotic vesicle pinches off from the plasma membrane during receptor-mediated endocytosis, the clathrin coat is removed, and clathrin proteins are recycled back to the membrane for reuse. The resulting vesicles exhibit several possible fates, depending upon cell type and the specific ligand engulfed. Some vesicles fuse with intracellular organelle membranes, adding their contents to that organelle's internal lumen. Other endocytotic vesicles traverse the cytoplasm and fuse with the plasma membrane on the opposite cell side, releasing their contents into the extracellular space. This transcellular pathway enables transfer of large molecules, such as proteins, across cellular layers separating two fluid compartments in the body—for example, between blood and interstitial fluid. A similar mechanism permits small amounts of macromolecules to cross the intestinal epithelium.
Most endocytotic vesicles fuse with a series of intracellular vesicles and tubular elements known as endosomes (Chapter 3), which are positioned between the plasma membrane and the Golgi apparatus. Like the Golgi apparatus, endosomes perform essential sorting functions, distributing vesicle contents and membrane components to various cellular locations. Some endocytotic vesicle contents are passed from endosomes to the Golgi apparatus, where ligands undergo modification and processing. Other vesicles fuse with lysosomes—organelles containing digestive enzymes that break down large molecules including proteins, polysaccharides, and nucleic acids. Fusion of endosomal vesicles with the lysosomal membrane exposes vesicle contents to these powerful digestive enzymes. Finally, in many cases, receptors internalized with the vesicle are recycled back to the plasma membrane for additional rounds of endocytosis.
Potocytosis. Another specialized fate of endocytotic vesicles occurs in a distinct form of receptor-mediated endocytosis called potocytosis. Potocytosis resembles other receptor-mediated endocytosis types in that an extracellular ligand typically binds to a plasma membrane receptor, initiating intracellular vesicle formation. However, in potocytosis, ligands appear primarily restricted to low-molecular-weight molecules such as certain vitamins, although lipoprotein complexes have also been observed to utilize this pathway. Potocytosis differs from clathrin-dependent receptor-mediated endocytosis primarily in the fate of the endocytotic vesicle. In potocytosis, tiny vesicles called caveolae (singular, caveola, meaning "little cave") pinch off from the plasma membrane and deliver their contents directly to the cell cytosol rather than merging with lysosomes or other organelles. Small molecules within caveolae may diffuse into the cytosol via channels or be transported by carrier proteins. Although their functions remain under active investigation, caveolae have been implicated in diverse important cellular functions, including cell signaling, transcellular transport, and cholesterol homeostasis.
Membrane Recycling During Endocytosis. Each endocytosis episode removes a small portion of membrane from the cell surface. In cells exhibiting high endocytotic activity, more than 100% of the plasma membrane may be internalized within a single hour, yet the membrane surface area remains remarkably constant. This stability occurs because membrane is replaced at approximately the same rate by vesicle membrane fusing with the plasma membrane during exocytosis. Some plasma membrane proteins internalized during endocytosis are stored within endosome membranes and, upon receiving appropriate signals, can be returned to fuse with the plasma membrane during exocytosis, completing the recycling loop.
Exocytosis. Exocytosis performs two essential functions for cells. First, it provides a mechanism to replace plasma membrane portions removed by endocytosis and, in doing so, offers a route for adding new membrane components. Second, exocytosis provides a pathway by which membrane-impermeable molecules (such as protein hormones) synthesized by the cell can be secreted into the extracellular fluid, enabling intercellular communication.
How do cells package substances destined for exocytotic secretion into vesicles? Chapter 3 described the entry of newly formed proteins into the endoplasmic reticulum lumen and subsequent protein processing through the Golgi apparatus. From the Golgi apparatus, proteins destined for secretion travel to the plasma membrane within vesicles, from which they can be released into the extracellular fluid by exocytosis. In some cases, substances enter vesicles via mediated transporters located in the vesicle membrane, adding another layer of regulation to the secretory pathway.
The secretion of substances by exocytosis is triggered in most cells by stimuli that elevate cytosolic Ca²⁺ concentration. As described in Chapters 5 and 6, these stimuli open calcium channels in the plasma membrane and/or intracellular organelle membranes. The resulting increase in cytosolic Ca²⁺ concentration activates proteins required for vesicle membrane fusion with the plasma membrane, releasing vesicle contents into the extracellular fluid. Material stored in secretory vesicles remains available for rapid secretion in response to stimuli, avoiding delays that would occur if synthesis were required only after stimulus arrival. Exocytosis represents the primary mechanism by which most neurons communicate through release of neurotransmitters stored in secretory vesicles that merge with the plasma membrane. This process also constitutes a major pathway for hormone release from endocrine cells into the extracellular fluid.
Cells that actively undergo exocytosis recover membrane bits through a process called compensatory endocytosis. This process—whose mechanisms remain uncertain but may involve both clathrin-mediated and non-clathrin-mediated events—restores membrane material to the cytoplasm that can be made available for forming new secretory vesicles. Compensatory endocytosis also helps prevent unchecked expansion of the plasma membrane, maintaining cellular surface area within physiological limits.
Date added: 2026-07-14; views: 3;