Epithelial Transport: Paracellular and Transcellular Pathways
As described in Chapter 1, epithelial cells line hollow organs or tubes and play a critical role in regulating the absorption or secretion of substances across these surfaces. One surface of an epithelial cell generally faces a hollow or fluid-filled tube or chamber, and the plasma membrane on this side is referred to as the apical membrane (also known as the luminal membrane; refer back to Figures 1.2 and 3.9). The plasma membrane on the opposite surface, which is usually adjacent to a network of blood vessels, is called the basolateral membrane (also known as the serosal membrane). These two membrane domains exhibit distinct compositions and functions, enabling directional transport across the epithelial layer.
Two primary pathways allow a substance to cross a layer of epithelial cells. The first is the paracellular pathway, in which diffusion occurs between adjacent epithelial cells through the intercellular space. The second is the transcellular pathway, where a substance moves into an epithelial cell across either the apical or basolateral membrane, diffuses through the cytosol, and exits across the opposite membrane (Figure 4.22). Diffusion via the paracellular pathway is significantly limited by the presence of tight junctions between adjacent cells, because these junctions form a continuous seal around the apical end of each epithelial cell (Chapter 3). Although small ions and water can diffuse to some degree through tight junctions, the overall extent of paracellular diffusion is constrained by both the tightness of the junctional seal and the relatively small surface area available for this route.
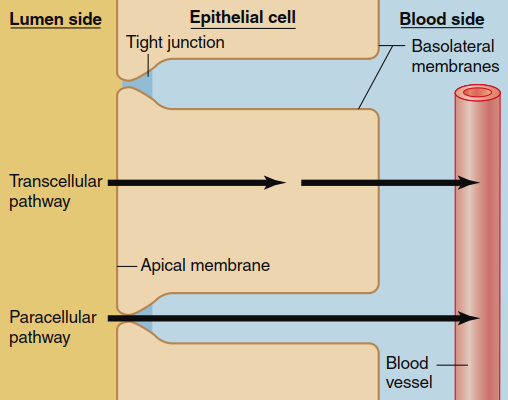
Figure 4.22. The two major routes by which water and solutes move across an epithelium, shown here as moving from the lumen of a tube or hollow chamber into the blood
During transcellular transport, the movement of molecules through the plasma membranes of epithelial cells occurs via the previously described mechanisms of diffusion and mediated transport. However, the transport and permeability characteristics of the apical and basolateral membranes are not identical; these two membrane domains often contain different ion channels and different transporters for mediated transport. As a consequence of these differences, substances can undergo net movement from a low concentration on one side of an epithelium to a higher concentration on the opposite side. Classic examples include the absorption of nutrients from the gastrointestinal tract into the blood, the movement of substances between kidney tubules and blood during urine formation, and the secretion of ions and water by glands such as sweat glands.
Figure 4.23 and Figure 4.24 illustrate two distinct examples of active transport across an epithelium. Sodium ions (Na⁺) are actively transported across most epithelia from the luminal side to the blood side. In the example shown in Figure 4.23, the movement of Na⁺ from the lumen into the epithelial cell occurs by diffusion through Na⁺ channels located in the apical membrane. Na⁺ diffuses into the cell because the intracellular Na⁺ concentration is maintained at a low level by the active transport of Na⁺ back out of the cell across the basolateral membrane, where all of the Na⁺/K⁺-ATPase pumps are exclusively located. In other words, Na⁺ moves downhill (passively) into the cell and then uphill (actively) out of it. The net result is that Na⁺ can be moved via the transcellular pathway from a region of lower concentration to a region of higher concentration across the entire epithelium, a process known as transepithelial transport.
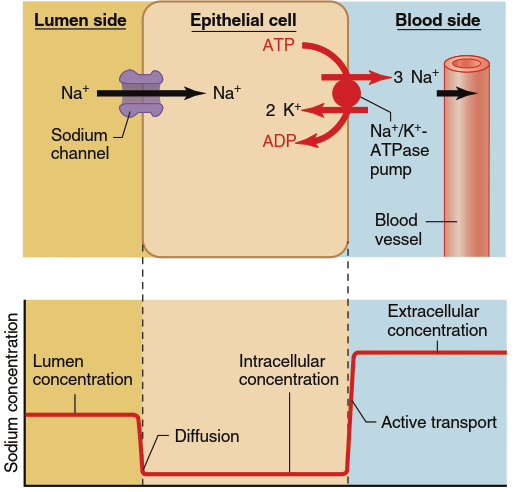
Figure 4.23. Active transport of Na+ across an epithelial cell. The transepithelial transport of Na+ always involves primary active transport out of the cell across one of the plasma membranes, typically via an Na+/K+-ATPase pump as shown here. The movement of Na+ into the cell across the plasma membrane on the opposite side is always downhill. Sometimes, as in this example, it is by diffusion through Na+ channels, whereas in other epithelia this downhill movement occurs through a secondary active transporter. Shown below the cell is the concentration profile of the transported solute across the epithelium
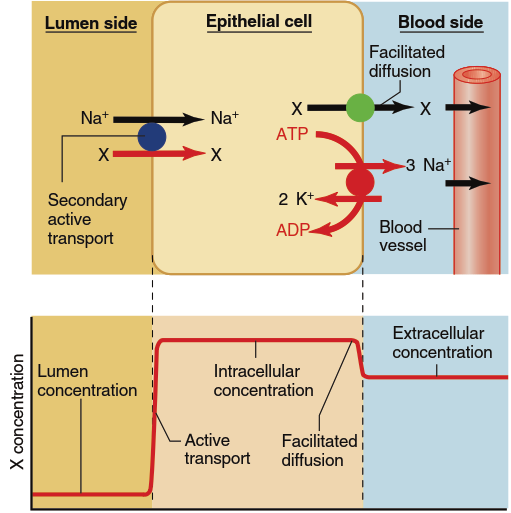
Figure 4.24. The transepithelial transport of most organic solutes (X) involves their movement into a cell through a secondary active transport driven by the downhill flow of Na+. The organic substance then moves out of the cell at the blood side down a concentration gradient by means of facilitated diffusion. Shown below the cell is the concentration profile of the transported solute across the epithelium
Figure 4.24 illustrates the active absorption of organic molecules across an epithelium, again via a transcellular pathway. In this case, the entry of an organic molecule (designated X) across the apical plasma membrane occurs via a secondary active transporter coupled to the downhill movement of Na⁺ into the cell. During this process, molecule X moves from a lower concentration in the luminal fluid to a higher concentration inside the cell, driven by the energy stored in the Na⁺ electrochemical gradient. The substance then exits across the basolateral membrane by facilitated diffusion, which moves the material from its higher intracellular concentration to a lower concentration in the extracellular fluid on the blood side. As a result, the concentration of the substance may become considerably higher on the blood side than in the lumen because the blood-side concentration can approach equilibrium with the high intracellular concentration created by the apical membrane entry step.
Although water is not actively transported across cell membranes, net movement of water across an epithelium can occur by osmosis as a direct consequence of the active transport of solutes, notably Na⁺, across the epithelium. The active transport of Na⁺, as previously described, produces a decrease in Na⁺ concentration on one side of an epithelial layer (the luminal side in our example) and an increase on the opposite (blood) side. These changes in solute concentration are accompanied by reciprocal changes in water concentration on the two sides because, as we have seen, a change in solute concentration inevitably produces a change in water concentration. The resulting water concentration difference will drive water movement by osmosis from the low-Na⁺ (high-water concentration) side to the high-Na⁺ (low-water concentration) side of the epithelium (Figure 4.25). Therefore, net movement of solute across an epithelium is accompanied by a flow of water in the same direction.
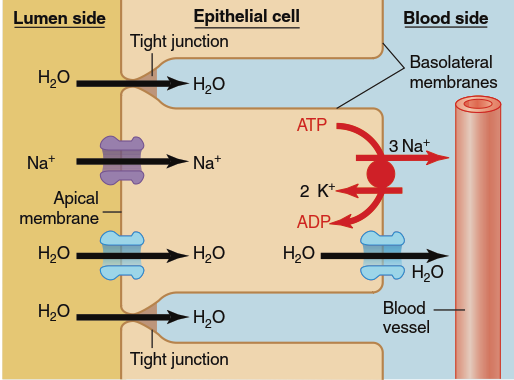
Figure 4.25. Net movements of water across an epithelium are dependent on net solute movements. The active transport of Na+ across the cells and into the surrounding interstitial spaces produces an elevated osmolarity in this region and a decreased osmolarity in the lumen. This leads to the osmotic flow of water across the epithelium in the same direction as the net solute movement. The water diffuses through aquaporins in the membrane (transcellular pathway) and across the tight junctions between the epithelial cells (paracellular pathway)
This principle has profound physiological implications. As you will learn in Chapter 14, this is a major mechanism by which epithelial cells of the kidney absorb water from the urine back into the blood, thereby concentrating urine and conserving body water. It is also the primary way in which water is absorbed from the intestines into the blood (Chapter 15), preventing dehydration and maintaining fluid balance. Additionally, similar mechanisms operate in salivary glands, pancreatic ducts, and sweat glands, where solute transport drives water secretion or absorption depending on the direction of the ion gradients.
Regulation of Epithelial Transport – It is important to note that epithelial transport is not static but highly regulated by hormones, neurotransmitters, and local factors. For example, aldosterone increases the number of Na⁺ channels and Na⁺/K⁺-ATPase pumps in kidney epithelial cells, enhancing Na⁺ reabsorption. Similarly, antidiuretic hormone (ADH) regulates water permeability by inserting aquaporin water channels into the apical membrane of collecting duct cells. Such regulatory mechanisms ensure that epithelial tissues can adapt to changing physiological conditions, maintaining homeostasis of fluids and electrolytes throughout the body.
Date added: 2026-07-14; views: 4;