Mechanisms of Mediated Transport: Transporters, Channels, and Cellular Physiology
A fundamental principle of physiology is the controlled exchange of materials between biological compartments and across cellular membranes. While simple diffusion and movement through gated ion channels account for some transmembrane movement, they cannot explain the passage of all substances. Many essential molecules, such as amino acids and glucose, are too polar to diffuse through the lipid bilayer and too large for channels. The movement of these molecules, along with non-diffusional ion movement, is facilitated by integral membrane proteins known as transporters. This process is termed mediated transport, which relies on specific conformational changes within these transporter proteins, as illustrated in the mechanism shown in Figure 4.8.
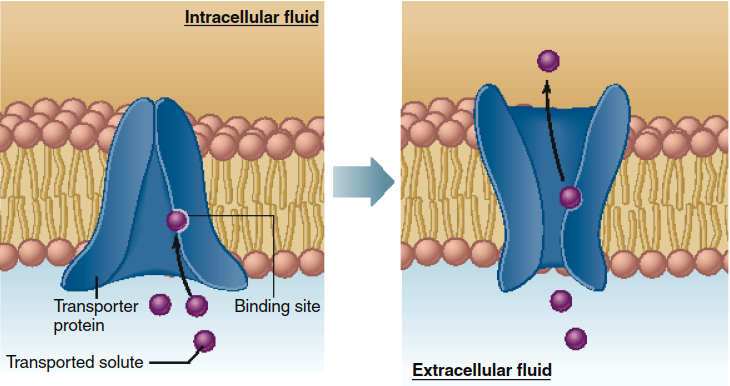
Figure 4.8. Model of mediated transport. A change in the conformation of the transporter exposes the transporter binding site first to one surface of the membrane then to the other, thereby transferring the bound solute from one side of the membrane to the other. This model shows net mediated transport from the extracellular fluid to the inside of the cell. In many cases, the net transport is in the opposite direction. The size of the conformational change is exaggerated for illustrative purposes in this and subsequent figures.
The process of mediated transport begins when the solute binds to a specific site on the transporter, a site initially exposed to one surface of the membrane. Following binding, a conformational change in the transporter protein exposes the binding site to the solution on the opposite side of the membrane. The subsequent dissociation of the solute from the transporter completes its transmembrane passage. This mechanism allows bidirectional movement, with molecules binding on one side and releasing on the other. It is important to note that the diagram in Figure 4.8 represents a model, as the precise conformational changes for most transport proteins remain an area of active research.
Transporters share several characteristics with ion channels, including their nature as membrane proteins and their property of chemical specificity. Their critical difference lies in transport capacity. Ion channels typically move ions several thousand times faster per unit time than transporters move molecules. This disparity exists because a transporter must undergo a conformational cycle for each molecule it moves, whereas an open ion channel permits a continuous passive flow of ions. An analogy is the difference in throughput between a continuous-flow bridge and a shuttle ferry.
Membranes contain many distinct types of transporters, each exhibiting specificity for a particular substance or class of related substances. For instance, a transporter for amino acids will not transport sugars, and vice versa. The plasma membranes of different cell types contain unique complements of transporters, leading to cell-specific differences in the types and rates of substance transport. This specialization is crucial for differentiated cellular function.
The magnitude of solute flux through a mediated-transport system is governed by four key factors: (1) the extracellular solute concentration, (2) the affinity of the transporters' binding sites for the solute, (3) the number of functional transporters in the membrane, and (4) the rate of the transporter's conformational change. Modulating any of these factors can regulate the transport flux. A central concept is transporter saturation. Since the number of transporters in a membrane is finite, as solute concentration increases, binding sites become occupied until they are saturated. At this point, the maximal transport flux (Vmax) is reached, and further increases in concentration yield no increased flux.
This saturation kinetics produces a flux curve that differs markedly from simple diffusion through the lipid bilayer. As shown in Figure 4.9, diffusion flux increases linearly with concentration, while mediated transport flux reaches a plateau at saturation. When saturated, the maximal transport rate is limited by the speed of the transporter's conformational cycle, which is inherently slower than ion flux through channels. This distinction underscores the different physiological roles of channels (rapid signaling) and transporters (controlled solute transfer).
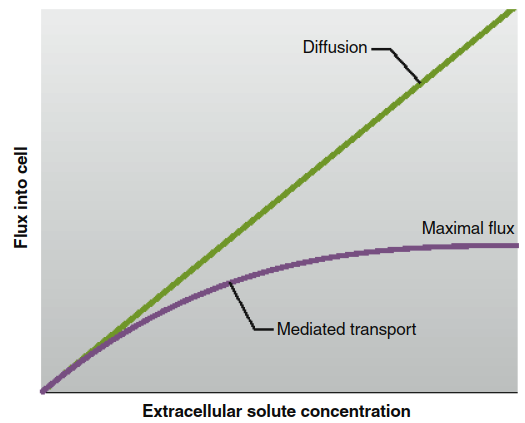
Figure 4.9. The flux of molecules diffusing into a cell across the lipid bilayer of a plasma membrane (green line) increases continuously in proportion to the extracellular concentration, whereas the flux of molecules through a mediated-transport system (purple line) reaches a maximal value.
Mediated transport systems are broadly categorized into two types: facilitated diffusion and active transport. In facilitated diffusion, the net movement of a solute is always down its concentration gradient, from higher to lower concentration. The key distinction from simple diffusion is the requirement for a specific transporter protein, like the model in Figure 4.8. This process does not require direct cellular energy input (ATP) and cannot move a solute against its gradient. Equilibrium is reached when concentrations equalize on both membrane sides.
A paramount example of facilitated diffusion is the transport of glucose by GLUT proteins (glucose transporters). These transporters are essential because the polar glucose molecule cannot efficiently cross the lipid bilayer on its own. Intracellular glucose concentration often remains lower than extracellular levels because incoming glucose is rapidly phosphorylated to glucose-6-phosphate for metabolism. This maintains a continuous inward diffusion gradient. Hormonal regulation of GLUT numbers in the plasma membrane allows the body to control metabolic substrate entry, a vital aspect of homeostasis.
In contrast, active transport moves a solute against its concentration gradient (uphill), which requires an energy input. Like facilitated diffusion, it involves specific solute binding to a transporter, exhibits specificity and saturation, and uses transporter proteins often called pumps. The maintenance of a steady-state concentration difference is energy-dependent. Energy coupling occurs via two main mechanisms: primary active transport and secondary active transport, distinguished by their immediate energy source.
Primary active transport directly utilizes the energy from ATP hydrolysis. The transporter itself is an ATPase enzyme that phosphorylates itself, inducing a conformational change that drives solute movement. The quintessential example is the Na+/K+-ATPase pump. This ubiquitous transporter moves three sodium ions (Na+) out of the cell and two potassium ions (K+) in per ATP hydrolyzed, against their respective gradients. The sequence, detailed in Figure 4.11, involves cyclical binding, phosphorylation, conformational change, ion release, dephosphorylation, and reset. This pump is fundamental to maintaining the characteristic high intracellular K+ and low intracellular Na+ concentrations (Figure 4.12) and contributes to the resting membrane potential.
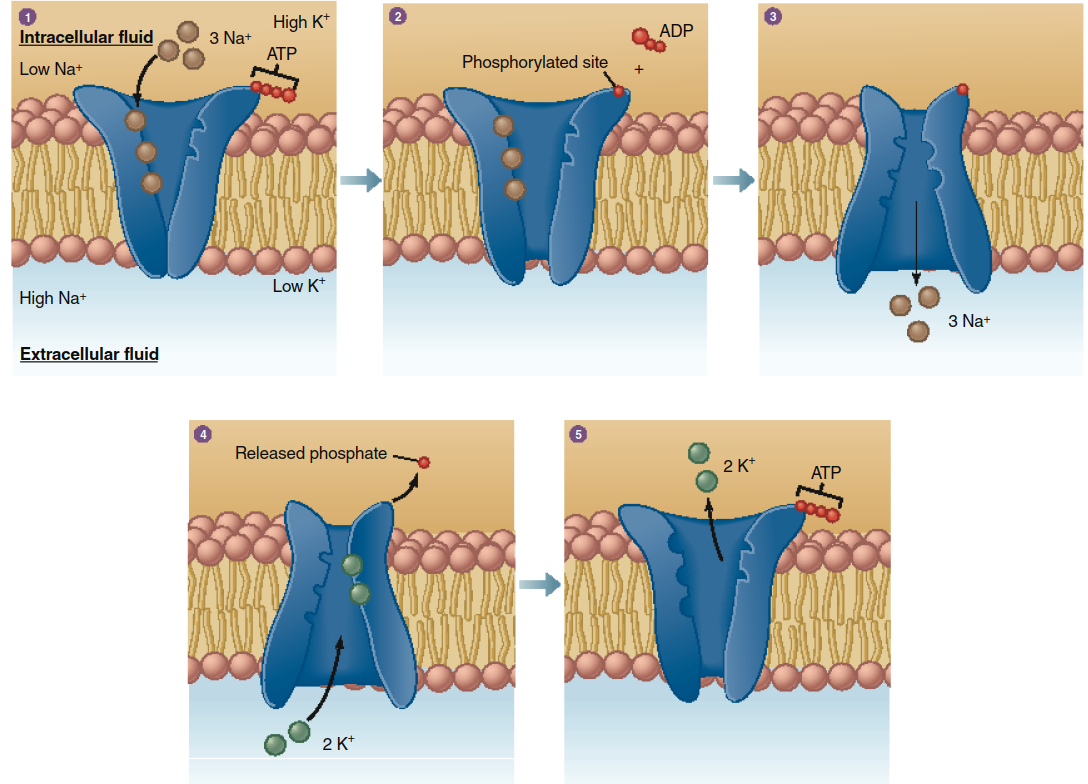
Figure 4.11. Active transport of Na+ and K+ mediated by the Na+/K+-ATPase pump. See text for the numbered sequence of events occurring during transport.
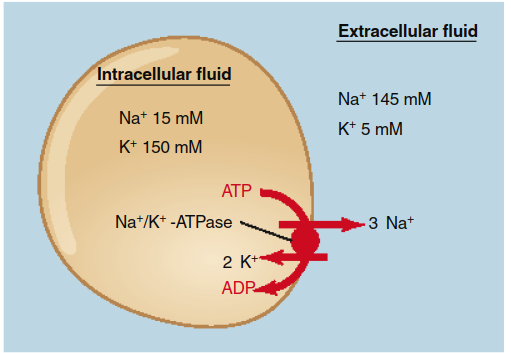
Figure 4.12. The primary active transport of sodium and potassium ions in opposite directions by the Na+/K+-ATPase in plasma membranes is responsible for the low Na+ and high K+ intracellular concentrations. For each ATP hydrolyzed, three sodium ions move out of a cell and two potassium ions move in.
Other critical primary active transporters include the Ca2+-ATPase, which maintains low cytosolic calcium by pumping Ca2+ into the extracellular fluid or into organelles like the endoplasmic reticulum; the H+-ATPase, which regulates cellular pH; and the H+/K+-ATPase, vital for stomach acid secretion. The collective activity of these pumps consumes a significant portion of the body's ATP, underscoring their physiological importance.
Secondary active transport uses the energy stored in an electrochemical gradient, typically the Na+ gradient established by the Na+/K+-ATPase, to drive the uphill transport of another solute. Transporters in this class have two binding sites: one for the ion moving downhill (usually Na+) and one for the co-transported solute moving uphill. As diagrammed in Figure 4.13, Na+ binding increases the affinity for the second solute. The conformational change driven by Na+ entry releases both into the cytosol. The Na+ gradient is subsequently restored by primary active transport, indirectly coupling secondary transport to metabolic energy.
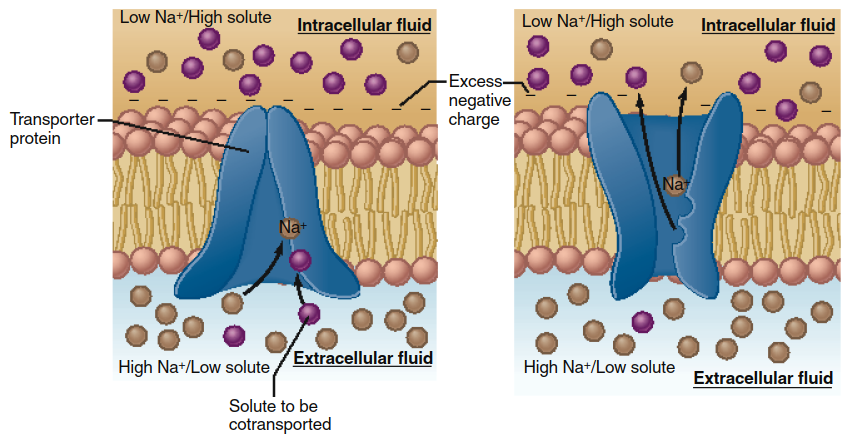
Figure 4.13. Secondary active-transport model. In this example, the binding of a sodium ion to the transporter produces an allosteric increase in the affinity of the solute binding site at the extracellular surface of the membrane. Binding of Na+ and solute causes a conformational change in the transporter that exposes the binding sites to the intracellular fluid. Na+ diffuses down its electrochemical gradient into the cell, which returns the solute binding site to a low-affinity state.
The direction of the second solute relative to Na+ defines two subtypes: cotransport (symport), where both solutes move in the same direction (e.g., Na+ and glucose into intestinal cells), and countertransport (antiport), where they move in opposite directions (e.g., Na+ in and Ca2+ or H+ out). These processes are illustrated in Figure 4.14. Ultimately, all secondary active transport derives its energy from ATP via the primary active transport that creates the ion gradient.
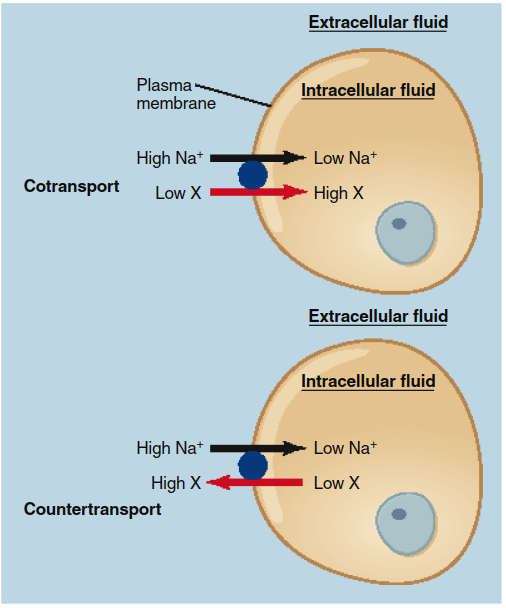
Figure 4.14. Cotransport and countertransport during secondary active transport driven by Na+. Sodium ions always move down their concentration gradient into a cell, and the transported solute always moves up its gradient. Both Na+ and the transported solute X move in the same direction during cotransport, but in opposite directions during countertransport.
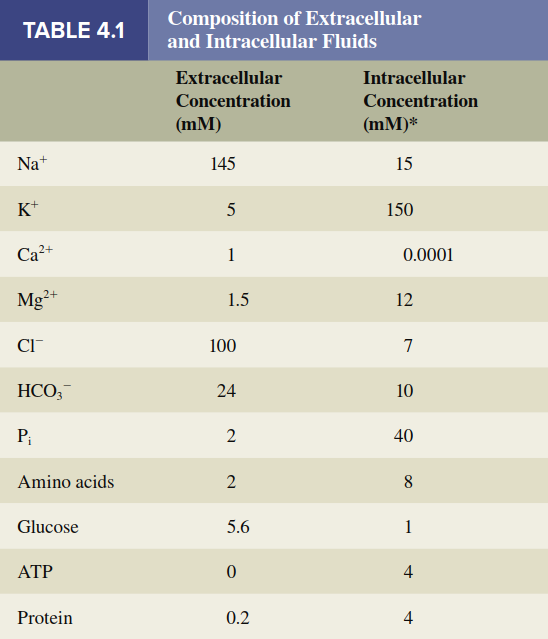
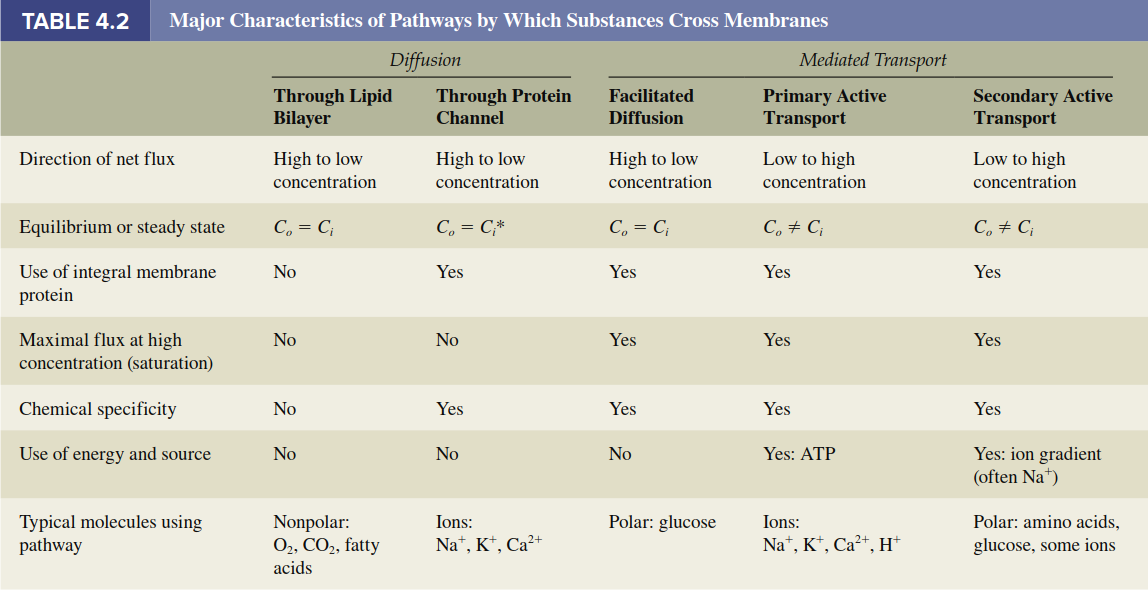
In summary, the unequal distribution of substances between intracellular and extracellular fluids (Table 4.1) results from the integrated actions of ion channels, the membrane potential, and primary and secondary active transporters. Table 4.2 provides a comparative summary of all major transmembrane movement pathways, and Figure 4.15 integrates these mechanisms schematically on a plasma membrane. A related but distinct process is the movement of water across membranes, a special case governed by osmosis, which will be discussed subsequently.
Date added: 2026-07-14; views: 4;