Signal Transduction Pathways: How Chemical Messengers Trigger Cellular Responses
The binding of a chemical messenger to its receptor causes a conformational change (tertiary structure alteration) in the receptor—an event known as receptor activation. This activation represents the initial step leading to the cell’s responses to the messenger. Cellular responses can take the form of changes in (1) the permeability, transport properties, or electrical state of the plasma membrane; (2) metabolism; (3) secretory activity; (4) rate of proliferation and differentiation; or (5) contractile or other activities.
Despite the variety of responses, a common denominator exists: all responses are directly due to alterations of specific cell proteins. For example, neurotransmitter-induced generation of electrical signals in neurons reflects the altered conformation of membrane proteins (ion channels) through which ions diffuse. Similarly, changes in the rate of glucose secretion by the liver induced by the hormone epinephrine reflect altered activity and concentration of enzymes in glucose synthesis pathways. Muscle contraction induced by the neurotransmitter acetylcholine results from the altered conformation of contractile proteins.
Receptor activation by a messenger is only the first step leading to the cell’s ultimate response (contraction, secretion, etc.). The diverse sequences of events that link receptor activation to cellular responses are termed signal transduction pathways. “Pathways” denotes the cell-specific mechanisms linked with different messengers. Signal transduction pathways differ fundamentally between lipid-soluble and water-soluble messengers, because their receptors reside in different locations—intracellular for lipid-soluble messengers and in the plasma membrane for water-soluble messengers.
Pathways Initiated by Lipid-Soluble Messengers. Lipid-soluble messengers include hydrophobic substances such as steroid hormones and thyroid hormone. Their receptors belong to a large family of intracellular receptors called nuclear receptors that share similar structures (see Figure 5.1b) and mechanisms of action. Although plasma membrane receptors for a few of these messengers have been identified, most receptors in this family are intracellular. In some cases, inactive receptors are located in the cytosol and move into the nucleus after binding their ligand; however, most inactive receptors already reside in the cell nucleus, where they bind to and are activated by their respective ligands. In both cases, receptor activation leads to altered rates of transcription of one or more genes in a particular cell.
In the most common scenario, the messenger diffuses out of capillaries from plasma to the interstitial fluid (refer back to Figure 1.3). From there, the messenger diffuses across the lipid bilayers of the plasma membrane and nuclear envelope to enter the nucleus and bind to the receptor there (Figure 5.4). The activated receptor complex then functions in the nucleus as a transcription factor—a regulatory protein that directly influences gene transcription. The hormone-receptor complex binds to DNA at a regulatory region of a gene, typically increasing the rate of that gene’s transcription into mRNA. The mRNA molecules move out of the nucleus to direct the synthesis, on ribosomes, of the protein the gene encodes. The result is an increase in the cellular concentration of the protein and/or its rate of secretion, accounting for the cell’s ultimate response to the messenger. For example, if the protein encoded by the gene is an enzyme, the cell’s response is an increase in the rate of the reaction catalyzed by that enzyme.
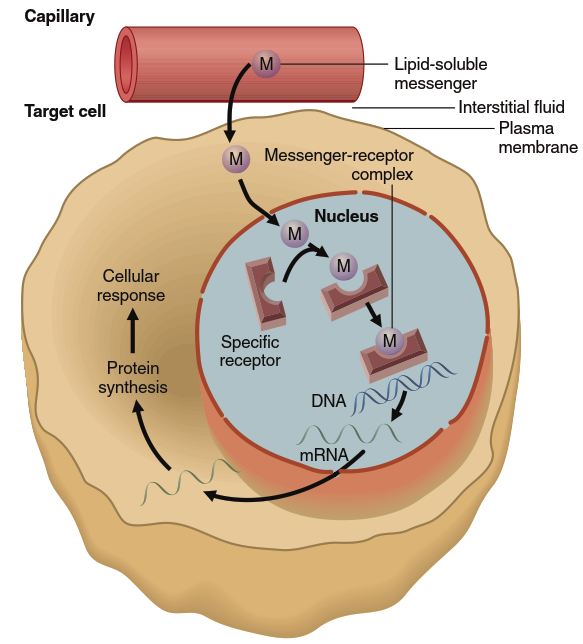
Figure 5.4. Mechanism of action of lipid-soluble messengers. This figure shows the receptor (simplified in this view) for these messengers in the nucleus. In some cases, the unbound receptor is in the cytosol rather than the nucleus, in which case the binding occurs there, and the activated messenger-receptor complex then moves into the nucleus. For simplicity, a single messenger is shown binding to a single receptor. In many cases, however, two messenger-receptor complexes must bind together in order to activate a gene
Two other points are important. First, more than one gene may be subject to control by a single receptor type. For example, the adrenal gland hormone cortisol acts via its intracellular receptor to activate numerous genes involved in the coordinated control of cellular metabolism and energy balance. Second, in some cases, the transcription of a gene or genes may be decreased rather than increased by the activated receptor. Cortisol inhibits transcription of several genes whose protein products mediate inflammatory responses following injury or infection; for this reason, cortisol has important anti-inflammatory effects.
Pathways Initiated by Water-Soluble Messengers. Water-soluble messengers cannot readily enter cells by diffusion through the lipid bilayer of the plasma membrane. Instead, they exert their actions by binding to the extracellular portion of receptor proteins embedded in the plasma membrane. Water-soluble messengers include most polypeptide hormones, neurotransmitters, and paracrine and autocrine compounds. The signal transduction mechanisms initiated by water-soluble messengers can be classified into four types illustrated in Figure 5.5.
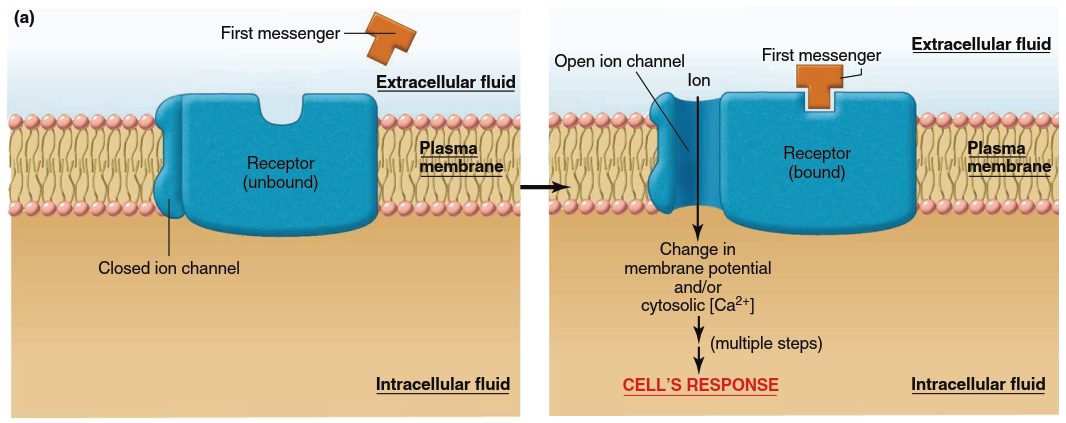
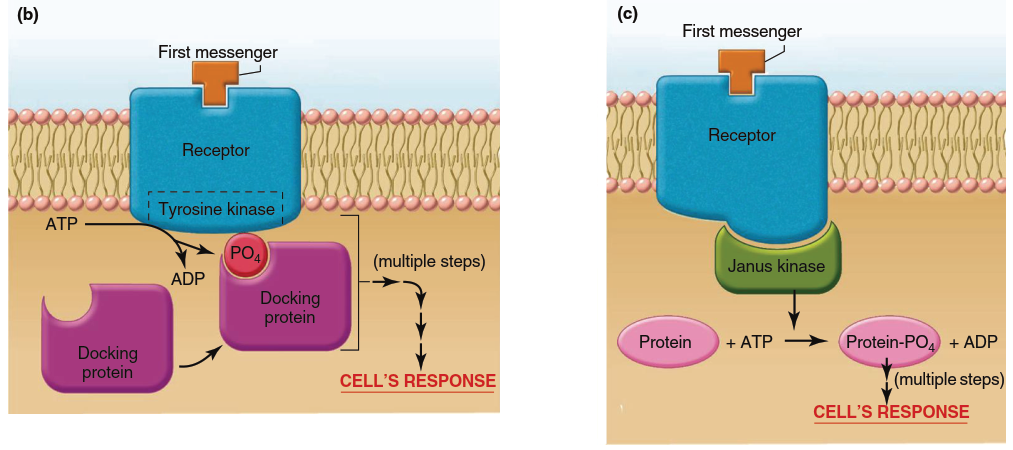
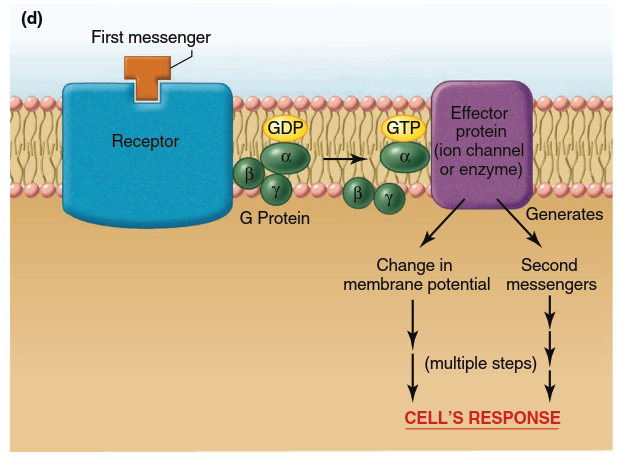
Figure 5.5. Mechanisms of action of water-soluble messengers (noted as “first messengers” in this and subsequent figures). (a) Signal transduction mechanism in which the receptor complex includes an ion channel. Note that the receptor exists in two conformations in the unbound and bound states. It is the binding of the first messenger to its receptor that triggers the conformational change that leads to opening of the channel. Note: Conformational changes also occur in panels b–d but only the bound state is shown for simplicity. (b) Signal transduction mechanism in which the receptor itself functions as an enzyme, usually a tyrosine kinase. (c) Signal transduction mechanism in which the receptor activates a janus kinase in the cytoplasm. (d) Signal transduction mechanism involving G proteins. When GDP is bound to the alpha subunit of the G protein, the protein exists as an inactive trimeric molecule. Binding of GTP to the alpha subunit causes dissociation of the alpha subunit, which then activates the effector protein
Three general terms are essential for this discussion. First, the extracellular chemical messengers (hormones or neurotransmitters) that reach the cell and bind to their specific plasma membrane receptors are often referred to as first messengers. Second messengers are substances that enter or are generated in the cytoplasm as a result of receptor activation by the first messenger; they diffuse throughout the cell to serve as chemical relays from the plasma membrane to the biochemical machinery inside the cell. Third, a protein kinase is an enzyme that phosphorylates other proteins by transferring a phosphate group from ATP to them. Phosphorylation allosterically changes a protein’s tertiary structure and consequently alters its activity. Different proteins respond differently—some are activated, some are inactivated. Many different protein kinases exist, each able to phosphorylate only specific proteins, and they are involved in signal transduction pathways that may include cascades where one protein kinase activates another, ultimately phosphorylating key proteins such as transporters, metabolic enzymes, ion channels, and contractile proteins. Enzymes that do the reverse—protein phosphatases—dephosphorylate proteins and also participate in signal transduction, serving to stop a signal once a cell response has occurred.
Signaling by Receptors That Are Ligand-Gated Ion Channels. In one type of plasma membrane receptor for water-soluble messengers, the protein that acts as the receptor is also an ion channel (refer back to Figure 4.7). Activation of the receptor by a first messenger (the ligand) results in a conformational change such that it forms an open channel through the plasma membrane (Figure 5.5a). These channels are known as ligand-gated ion channels (described in Chapter 4) and are particularly prevalent in the plasma membranes of neurons and skeletal muscle (Chapters 6 and 9). The opening of ligand-gated ion channels in response to first messenger binding increases net diffusion across the plasma membrane of one or more types of ions specific to that channel. As introduced in Chapter 4 (see Figure 4.6), such a change in ion diffusion results in a change in the electrical charge, or membrane potential, of a cell—this change in membrane potential is the cell’s response to the messenger. Additionally, when the channel is a Ca²⁺ channel, its opening increases cytosolic Ca²⁺ concentration, another essential event in many signaling systems.
Signaling by Receptors That Function as Enzymes. Other plasma membrane receptors for water-soluble messengers have intrinsic enzyme activity. With one major exception, the many receptors that possess intrinsic enzyme activity are all protein kinases (Figure 5.5b). Of these, the great majority specifically phosphorylate tyrosine residues; consequently, these receptors are known as receptor tyrosine kinases. The typical sequence is as follows: binding of a specific messenger to the receptor changes its conformation so that its enzymatic portion (located on the cytoplasmic side) is activated, resulting in autophosphorylation of the receptor (the receptor phosphorylates some of its own tyrosine residues). The newly created phosphotyrosines on the cytoplasmic portion serve as docking sites for cytoplasmic proteins. The bound docking proteins then bind and activate other proteins, which in turn activate one or more signaling pathways within the cell. The common denominator is that these pathways all involve activation of cytoplasmic proteins by phosphorylation.
There is one physiologically important exception: a receptor that functions both as a receptor and as a guanylyl cyclase to catalyze the formation, in the cytoplasm, of cyclic GMP (cGMP). In turn, cGMP functions as a second messenger to activate cGMP-dependent protein kinase, which phosphorylates specific proteins that mediate the cell’s response. As described in Chapter 7, such receptors are abundantly expressed in the retina of the eye, where they process visual inputs. This pathway is used by only a small number of messengers. Also, in certain cells, guanylyl cyclase enzymes are present in the cytoplasm; the first messenger—the gas nitric oxide (NO)—diffuses into the cytosol and combines with guanylyl cyclase to trigger cGMP formation. Nitric oxide is a lipid-soluble gas produced from the amino acid arginine by the action of nitric oxide synthase, present in numerous cell types including those lining blood vessels. When released, NO acts locally in a paracrine fashion to relax smooth muscle in certain blood vessels, allowing vasodilation—an important part of homeostatic blood pressure control.
Signaling by Receptors That Interact with Cytoplasmic Janus Kinases. In this category of signal transduction mechanisms for water-soluble messengers (Figure 5.5c), the enzymatic activity—again, tyrosine kinase activity—resides not in the receptor but in a family of separate cytoplasmic kinases called janus kinases (JAKs) that are associated with the receptor. The receptor and its associated janus kinase function as a unit. Binding of a first messenger to the receptor causes a conformational change leading to activation of the janus kinase. Different receptors associate with different members of the janus kinase family, and the different janus kinases phosphorylate different target proteins, many of which act as transcription factors. The result of these pathways is the synthesis of new proteins that mediate the cell’s response to the first messenger. One significant example of signals mediated primarily via receptors linked to janus kinases are those of the cytokines—proteins secreted by cells of the immune system that have critical functions in immune defenses.
Signaling by G-Protein-Coupled Receptors. The fourth category of signaling pathways for water-soluble messengers is by far the largest, including hundreds of distinct receptors (Figure 5.5d). Bound to the inactive receptor is a protein complex on the cytosolic surface of the plasma membrane belonging to the family of G proteins. G proteins contain three subunits: alpha, beta, and gamma. The alpha subunit can bind GDP and GTP; the beta and gamma subunits help anchor the alpha subunit in the membrane. Binding of a first messenger to the receptor changes the receptor’s conformation, increasing the affinity of the alpha subunit for GTP. When bound to GTP, the alpha subunit dissociates from the beta and gamma subunits. This dissociation allows the activated alpha subunit to link up with another plasma membrane protein—either an ion channel or an enzyme—that serves as an effector protein mediating the next steps in the sequence leading to the cell’s response.
Thus, a G protein serves as a switch to couple a receptor to an ion channel or to an enzyme in the plasma membrane; consequently, these receptors are known as G-protein-coupled receptors. The G protein may cause the ion channel to open, resulting in changes in electrical signals or (in the case of Ca²⁺ channels) changes in cytosolic Ca²⁺ concentration. Alternatively, the G protein may activate or inhibit the membrane enzyme with which it interacts; such enzymes, when activated, cause the generation of second messengers inside the cell. Once the alpha subunit of the G protein activates its effector protein, a GTPase activity inherent in the alpha subunit cleaves GTP into GDP and Pi, rendering the alpha subunit inactive and allowing it to recombine with its beta and gamma subunits.
There are several subfamilies of plasma membrane G proteins, each with multiple distinct members, and a single receptor may be associated with more than one type of G protein. Moreover, some G proteins may couple to more than one type of plasma membrane effector protein. In this way, a first-messenger-activated receptor, via its G-protein couplings, can call into action a variety of plasma membrane proteins such as ion channels and enzymes, which in turn induce a variety of cellular events. To illustrate major points concerning G proteins, plasma membrane effector proteins, second messengers, and protein kinases, the two most common effector protein enzymes regulated by G proteins—adenylyl cyclase and phospholipase C—are described in subsequent sections.
Date added: 2026-07-14; views: 4;