ABA Signal Transduction. ABA-Independent Signalling
ABA biosynthesis is known to occur predominantly in vascular parenchyma cells of roots and shoots (Fig. 6.18). Both the responsible enzymes and the expression of the respective genes have been detected there. Plasma membrane-localised ABC-type transporters such as ABCG25 in A. thaliana can export ABA. Cells respond to ABA synthesised by the cell itself and to ABA taken up from the apoplast. Uptake of ABA into guard cells is mediated by another ABC-type transporter (ABCG40 in A. thaliana). Lack of its activity reduces the responsiveness of guard cells to ABA (Fig. 6.19).
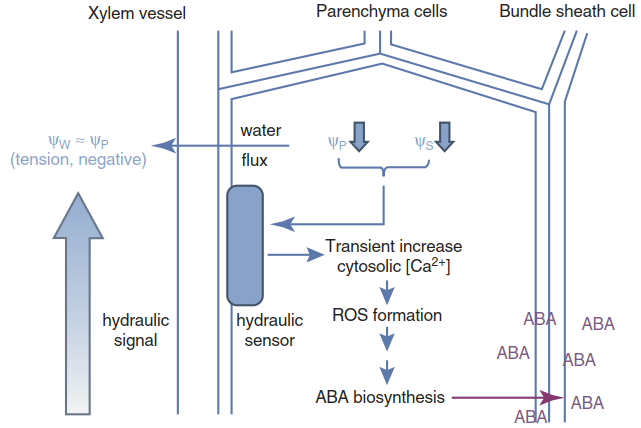
Fig. 6.18. Model of hydraulic signalling: a long-distance hydraulic signal (decrease in water potential (Ψw)) travels through the plant in the xylem and is locally converted into a biochemical signal. The decrease in Ψw causes water flux along the Ψw gradient out of the parenchyma cells within, for example, the shoot vasculature, as shown here. The resulting decrease in turgor pressure (Ψp) and osmotic potential (ΨS) is sensed by an unidentified receptor, which triggers a signalling cascade (a transient increase in cytosolic Ca2+ and production of reactive oxygen species (ROS)). The signalling activates ABA biosynthesis. ABA synthesised in parenchyma cells is exported to bundle sheath cells and beyond to trigger acclimative responses. (Modified from Christmann et al. (2013))
The earliest events in ABA signalling are mediated by a central regulatory module, which consists of three protein classes, the soluble ABA receptor PYR (PYRABACTIN RESISTANCE; also called RCAR, for REGULATORY COMPONENTS OF ABA RECEPTORS), protein phosphatases 2C (PP2C) and protein kinases (SnRK2s for SNF1-related protein kinases 2) (Fig. 6.20).
Several isoforms of each of these proteins are encoded by plant genomes. This provides flexibility for the ABA signalling, which occurs in all kinds of cells and during all developmental stages. The signalling pathway formed by the three types of proteins is double negative. In the inactive state the protein SnRK2, which triggers the ABA responses, is inhibited by PP2C. Upon binding of ABA the receptor PYR interacts with PP2C and thereby inactivates it. This releases SnRK2 from the inhibition and enables the phosphorylation of several possible target proteins.
The comparatively slow transcriptional responses to ABA are brought about by the phosphorylation of transcription factors such as AREB1 and AREB2. In the phosphorylated state these proteins are able to interact with ABA response elements (ABRE) in the promoters of ABA-responsive genes and to activate transcription. Target genes of these and related transcription factors encode, for instance, protective proteins such as LEA proteins and enzymes involved in compatible solute synthesis.
ABI5 is a transcription factor whose target genes maintain seed dormancy—one of the developmental processes under ABA control. Thus, the core signalling module is important for all types of ABA-dependent processes.
Rapid responses of guard cells are elicited by the activation of the anion channel SLAC1 and the inactivation of the inward-rectifying K+ channel KAT1. The resulting depolarisation of the plasma membrane opens outward-rectifying K+ channels, resulting in K+ efflux, turgor loss and stomatal closure (Figs. 6.13 and 6.14). SnRK2 also triggers additional signalling events such as the formation of ROS by NADPH oxidases (e.g. RbohF).
Besides ROS, other second messengers are involved in ABA signalling. Transient increases in cytosolic Ca2+ concentrations, as a very common element of signalling cascades (Chap. 2), are observed in ABA-treated guard cells too. The central ABA signalling module (Fig. 6.20) is not Ca2+ dependent. Instead, Ca2+ signalling modulates targets of the core pathway. ROS production stimulates a cytosolic Ca2+ increase which, via the action of Ca2+- dependent protein kinases, affects the activity of anion channels such as SLAC1. It is conceivable that the necessary integration of various environmental and physiological cues in guard cells, which is essential for a plant responding to many simultaneously changing aspects of its natural environment, is at least in part achieved by the convergence of several signalling pathways on the anion channels as central control points for the plasma membrane potential. Anion channel phosphorylation in multiple sites by different kinases controlled by distinct signalling pathways could provide a mechanism for such signal integration (Kollist et al. 2014).
ABA-Independent Signalling. ABA is clearly the central stress hormone controlling water deficit responses. Still, in mutant plants unable to respond to ABA, there can be induction of drought stress-responsive genes. This demonstrates the existence of signalling pathways that do not require ABA. A known pathway activates transcription factors of the CBF/DREB class, which are involved in ABA-independent signalling during cold acclimation. Some of these proteins trigger dehydration responses instead. The promoters of their target genes contain a cis element called the dehydration responsive element (DRE) or C-repeat.
Date added: 2025-01-27; views: 450;