Sensing of Water Status and Signal Transduction
As described above, a multitude of responses and acclimations is triggered by water deficit. Since the plant is part of the hydraulic soil-plant-air continuum and the cells are symplastically connected (with the exception of guard cells) (Steudle 2001), responses have to be coordinated in the entire organism. The stomatal aperture, for instance, has to be synchronised with the resistance of leaf and root tissue to water flow. Water shortage is translated into the stimulation of root growth. There is ample evidence for rapid responses of shoots to the root water status, and vice versa. Thus, long-distance signalling has to be in operation.
The most important endogenous signalling molecule mediating these responses is ABA. Its synthesis is rapidly and strongly up-regulated under drought stress (Fig. 6.17). Mutants lacking the ability to synthesise ABA are not able to withstand even a mild water deficit. ABA activates the majority of avoidance and tolerance mechanisms, including stomatal closure, the regulation of aquaporin abundance and activity, synthesis of protective proteins and osmoregulation.
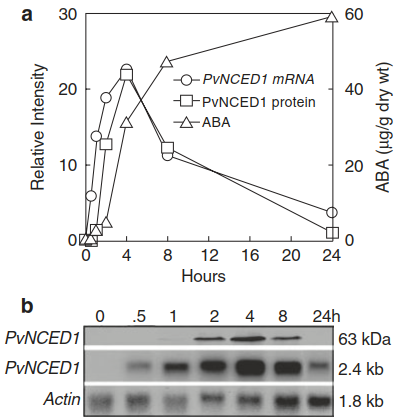
Fig. 6.17.Abscisic acid (ABA) synthesis is rapidly up-regulated upon dehydration of leaves. Upon water deficit, genes encoding the nine-cis-epoxycarotenoid dioxygenases (NCEDs) are transcriptionally activated. NCEDs catalyse the rate-limiting step in ABA synthesis. This activation results in a rapid rise in ABA concentrations. a Time course of changes in NCED messenger RNA (mRNA) and NCED protein abundance, as well as ABA concentrations occurring in detached, wilting Phaseolus vulgaris leaves. b Western blot and northern blot analysis of NCED protein and NCED mRNA, respectively. (Qin and Zeevaart 1999)
There is, however, also ABA-independent drought stress signalling. Furthermore, the rapid signalling of water status from the roots to the shoots, and vice versa, cannot be explained by ABA alone. ABA increases have frequently been detected in the xylem sap. However, grafting experiments with tomato and A. thaliana plants have demonstrated that leaves can respond normally to water deficit perceived by roots even when the roots are unable to synthesise ABA, so no ABA can travel to the shoot with the transpiration stream (Christmann et al. 2007).
ABA is an ancient molecule, detectable in all organisms with the exception of Archaea. Still, phylogenetic analysis of the main components of the core ABA signalling module (Fig. 6.20) suggests that ABA-dependent signalling apparently evolved in land plants (Hauser et al. 2011). Most drought stress avoidance and tolerance mechanisms are ABA dependent.
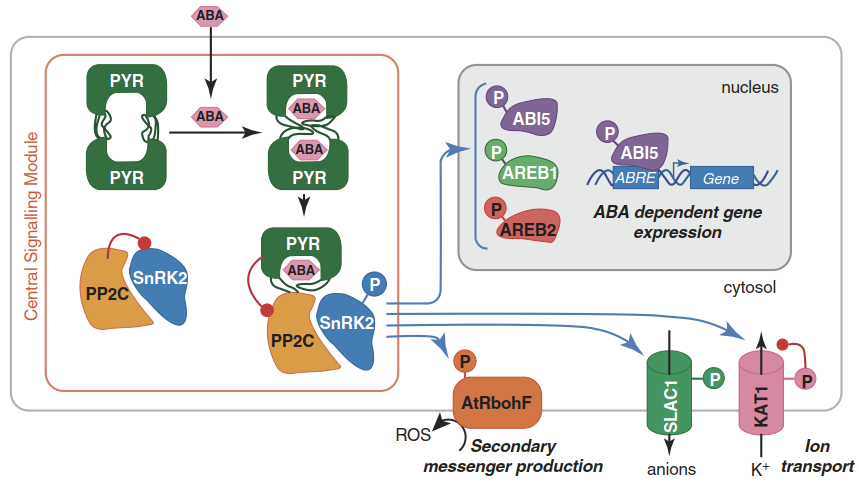
Fig. 6.20. The core abscisic acid (ABA) signalling module. Protein phosphatases 2C (PP2Cs, yellow) inhibit protein kinases (SnRKs) (indicated by the red symbol) in the absence of ABA. ABA is bound by intracellular receptors (PYR dimers), which dissociate and form ABA receptor- PP2C complexes. This complex formation inhibits PP2Cs and allows activation of SnRKs. SnRKs then phosphory- late (indicated by blue arrows) target proteins. Several SnRK targets are known, of which only a subset are depicted here. ABA can activate both fast and slow responses because the SnRKs can modulate either ion channel activity or transcription factor activity, respectively. In addition, SnRKs can trigger downstream signalling events—for instance, by activating reduced nicotinamide adenine dinucleotide phosphate (NADPH) oxidases (AtRbohF), which produce reactive oxygen species. (Hubbard et al. 2010)
Also, ABA triggers both rapid responses (within minutes), such as the closing of stomata, and responses that require changes in gene expression—that is, responses that depend on the interaction of transcription factors with cis elements in the promoters of stress tolerance genes and take hours to days. Three layers of ABA action can be differentiated: synthesis and transport, perception, and signal transduction (Hauser et al. 2011).
Date added: 2025-01-27; views: 447;