Evolution of Crassulacean Acid Metabolism Photosynthesis
The physiological plasticity of CAM corresponds to its evolutionary diversity. CAM manifestation is strongly influenced by the history of a species and its habitat context (Silvera et al. 2010). CAM is taxonomically more broadly distributed than C4 photosynthesis and is most likely evolutionary older. The presence of CAM in ancient groups such as the Isoetes (see I. howellii above) suggests a first appearance of CAM already in the Triassic.
Further CAM evolution was then probably driven by selection for increased carbon gain and better water use efficiency after the global reduction in atmospheric CO2 concentration about 30 million years ago during the Oligocene (see evolution of C4 photosynthesis, Sect. 6.6.2). CAM has evolved many times independently and to varying degrees. The extent of CAM manifestation shows a positive correlation with the dryness of the site; that is, the stronger the water scarcity of a habitat, the higher the likelihood of full CAM expression in the plants populating it.
Several characteristics of CAM can be distinguished, as described above (Borland et al. 2014). Accordingly, CAM requires a number of evolutionary changes in basal mechanisms that principally are present in all higher plants (Fig. 6.25):
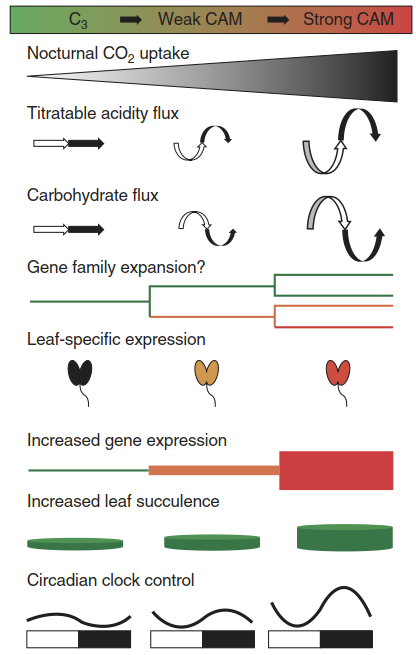
Fig. 6.25. Evolutionary changes required for crassulacean acid metabolism (CAM) photosynthesis. (After Silvera et al. (2010))
- First, a reversal of stomatal regulation enables nocturnal CO2 uptake. The control of the stomatal aperture by light has to be overridden by other control mechanisms. One factor could be the low internal leaf CO2 partial pressure at night, due to the activity of PEP carboxylase
- Second, diurnal fluctuations in organic acids and (reciprocally) in storage compounds and soluble sugars, plus respective transport activities, are established (e.g. malate into the vacuole and malic acid out of the vacuole)
- Third, key elements of CAM photosynthesis, e.g. carboanhydrase and PEPC, as well as decarboxylating enzymes—are more strongly expressed. Prerequisites here are the duplication and diversification of genes encoding the respective enzymes. As in plants with C4 photosynthesis, there are CAM plant-specific isoforms of PEP carboxylase with very high leaf expression
- Fourth, enhanced gluconeogenic and glycolytic activities supply substrates for carboxylation and decarboxylation
- The fifth element is leaf succulence. A clear correlation exists between the degree of leaf succulence and the strength of CAM. Plants with thicker leaves show lower δ13C values, which is indicative of stronger CAM (see the range of δ13C values in Table 6.3). This is explained not only by the greater storage capacity of larger cells (with vacuoles taking up 90-95% of the volume) but also by the tight packing of cells in succulent tissues, which restricts the intercellular gas space and thereby the gas exchange rates, and consequently limits C3 photosynthesis during phases II and IV
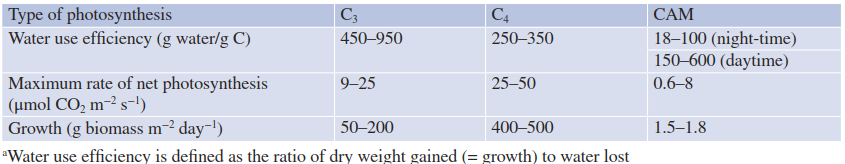
Table 6.3. Water use efficiency, photosynthesis and biomass production of C3, C4 and CAM plants. Crassulacean acid metabolism (CAM) plants are superior to other photosynthetic types in their water use efficiencya, but their photosynthetic rates and growth rates are much lower (Lüttge et al. 1994)
- Finally, the sixth key mechanism is the circadian clock control over CO2 fixation. Comparative studies with four Clusia species (one C3 species, two C3-CAM intermediates and one strong, constitutive CAM species) have revealed an association of the circadian control of PEPCK transcript abundance with CAM strength—that is, with day/ night changes in malate and soluble sugar content
As indicated in Fig. 6.25, all of these mechanisms can vary in their extent along a continuum of CAM manifestations.
References:
Agre P (2004) Aquaporin water channels (Nobel lecture). Angew Chem Int Ed Engl 43:4278-4290
Bartels D, Alexander R, Schneider K et al (1993) Desiccation-related gene products analyzed in a resurrection plant and in barley embryos. In: Close TJ, Bray EA (eds) Plant responses to cellular dehydration during environmental stress, Current topics in plant physiology, vol 10, pp 119-127. American Society of Plant Physiologists, Rockville
Borland AM, Hartwell J, Weston DJ et al (2014) Engineering crassulacean acid metabolism to improve water-use efficiency. Trends Plant Sci 19:327-338
Chater CCC, Oliver J, Casson S, Gray JE (2014) Putting the brakes on: abscisic acid as a central environmental regulator of stomatal development. New Phytol 202:376-391
Chaumont F, Tyerman SD (2014) Aquaporins: highly regulated channels controlling plant water relations. Plant Physiol 164:1600-1618
Christin P-A, Osborne CP (2014) The evolutionary ecology of C4 plants. New Phytol 204:765-781 Christmann A, Grill E, Huang J (2013) Hydraulic signals in long-distance signaling. Curr Opin Plant Biol 16:293-300
Christmann A, Weiler EW, Steudle E, Grill E (2007) A hydraulic signal in root-to-shoot signalling of water shortage. Plant J 52:167-174
Claeys H, Inze D (2013) The agony of choice: how plants balance growth and survival under water-limiting conditions. Plant Physiol 162:1768-1779
Cushman JC (2001) Crassulacean acid metabolism. A plastic photosynthetic adaptation to arid environments. Plant Physiol 127:1439-1448
Cushman JC, Bohnert HJ (1999) Crassulacean acid metabolism: molecular genetics. Annu Rev Plant Physiol Plant Mol Biol 50:305-332
Cushman JC, Tillett RL, Wood JA et al (2008) Large-scale mRNA expression profiling in the common ice plant, Mesembryanthemum crystallinum, performing C3 photosynthesis and crassulacean acid metabolism (CAM). J Exp Bot 59:1875-1894
Farrant JM, Moore JP (2011) Programming desiccation- tolerance: from plants to seeds to resurrection plants. Curr Opin Plant Biol 14:340-345
Gowik U, Westhoff P (2011) The path from C-3 to C-4 photosynthesis. Plant Physiol 155:56-63
Harb A, Krishnan A, Ambavaram MMR, Pereira A (2010) Molecular and physiological analysis of drought stress in Arabidopsis reveals early responses leading to acclimation in plant growth. Plant Physiol 154: 1254-1271
Date added: 2025-01-27; views: 557;