Neuron Structure, Axonal Transport, and Functional Classification
Neurons exhibit remarkable diversity in size and shape, yet they all share essential features enabling cell-to-cell communication. Long extensions known as processes connect neurons to one another and perform input and output functions. As illustrated in Figure 6.1, most neurons contain a cell body (soma) along with two types of processes: dendrites and axons. These structural components are fundamental to neural signaling and integration.
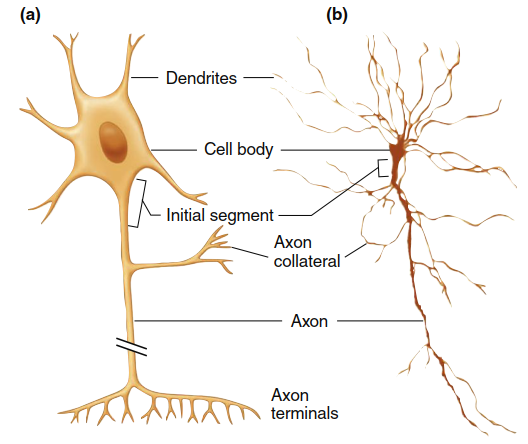
Figure 6.1. (a) Diagrammatic representation of one type of neuron. The break in the axon indicates that axons may extend for long distances; in fact, they may be 5000 to 10,000 times longer than the cell body is wide. This neuron is a common type, but there is a wide variety of neuronal morphologies, some of which have no axons.(b) A neuron as observed through a microscope. Dendritic spines and axon terminals cannot be seen at this magnification
The cell body (or soma) houses the nucleus and ribosomes, providing the genetic blueprint and machinery required for protein synthesis. Dendrites are highly branched outgrowths that receive incoming signals from other neurons. Their extensive branching increases the cell’s surface area—some central nervous system (CNS) neurons may possess up to 400,000 dendrites. Additionally, knoblike dendritic spines further expand the dendritic surface, enhancing the neuron’s capacity to receive inputs from numerous other cells.
The axon, also called a nerve fiber, is a long process extending from the cell body that carries outgoing signals to target cells. In humans, axons range from a few microns to over one meter in length. The region where the axon arises from the cell body is termed the initial segment (or axon hillock), and in most neurons this is where propagated electrical signals are generated. These signals then travel away from the cell body along the axon, which may give off branches known as collaterals. Near their ends, both the axon and its collaterals undergo further branching (see Figure 6.1); the greater the branching, the larger the neuron’s sphere of influence.
Each branch terminates in an axon terminal, responsible for releasing neurotransmitters from the axon. These chemical messengers diffuse across an extracellular gap to reach the opposing cell. Alternatively, some neurons release neurotransmitters from a series of bulging areas along the axon called varicosities. This process is critical for synaptic transmission and intercellular communication.
Many axons are covered by myelin sheaths (Figure 6.2), typically composed of 20 to 200 layers of highly modified plasma membrane wrapped around the axon by a nearby supporting cell. In the brain and spinal cord, these myelin-forming cells are a type of glial cell known as oligodendrocytes. A single oligodendrocyte may branch to form myelin on as many as 40 axons. In the peripheral nervous system (PNS), Schwann cells create individual myelin sheaths surrounding 1‑ to 1.5‑mm‑long segments at regular intervals along some axons. The gaps between adjacent myelin sections, where the axon’s plasma membrane is exposed to extracellular fluid, are called nodes of Ranvier. The myelin sheath accelerates the conduction of electrical signals along the axon and conserves energy.
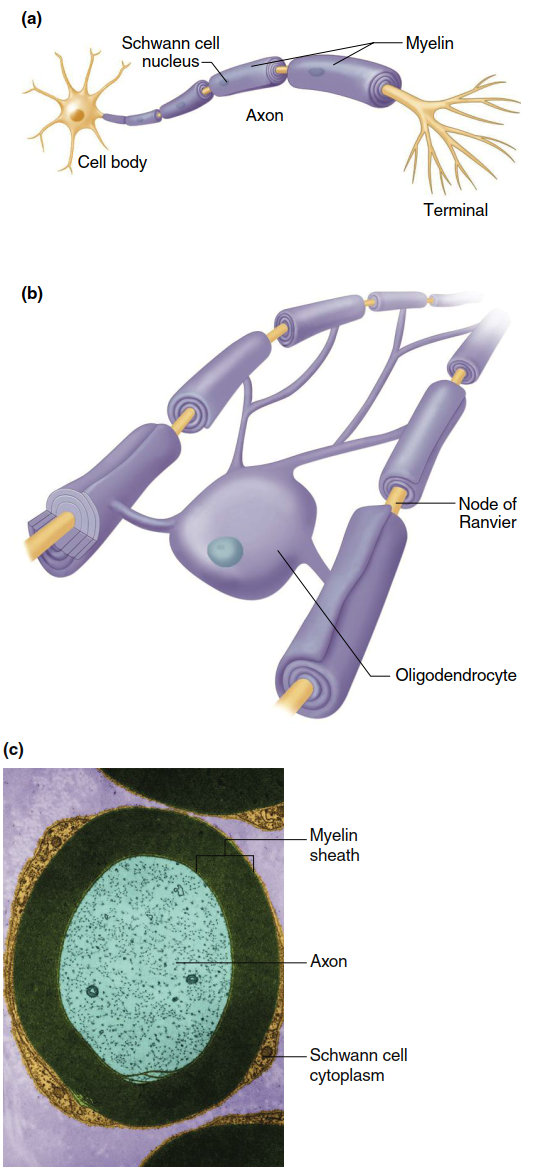
Figure 6.2. (a) Myelin formed by Schwann cells, and (b) oligodendrocytes on axons. (c) False color photomicrograph of a section through a myelinated axon in the PNS
To maintain axonal structure and function, various organelles and materials must travel up to one meter between the cell body and axon terminals. This movement, termed axonal transport, relies on a scaffolding of microtubule “rails” running the length of the axon and specialized motor proteins called kinesins and dyneins (Figure 6.3). These double‑headed motor proteins bind cellular cargo at one end and use energy from ATP hydrolysis to “walk” along microtubules. Kinesin primarily mediates transport from the cell body toward axon terminals (anterograde movement), delivering nutrient molecules, enzymes, mitochondria, neurotransmitter‑filled vesicles, and other organelles. Dynein moves in the opposite direction (retrograde), carrying recycled membrane vesicles, growth factors, and chemical signals that influence neuronal morphology, biochemistry, and connectivity. Retrograde transport also provides a route by which harmful agents such as tetanus toxin, herpes simplex virus, rabies virus, and poliovirus invade the CNS.
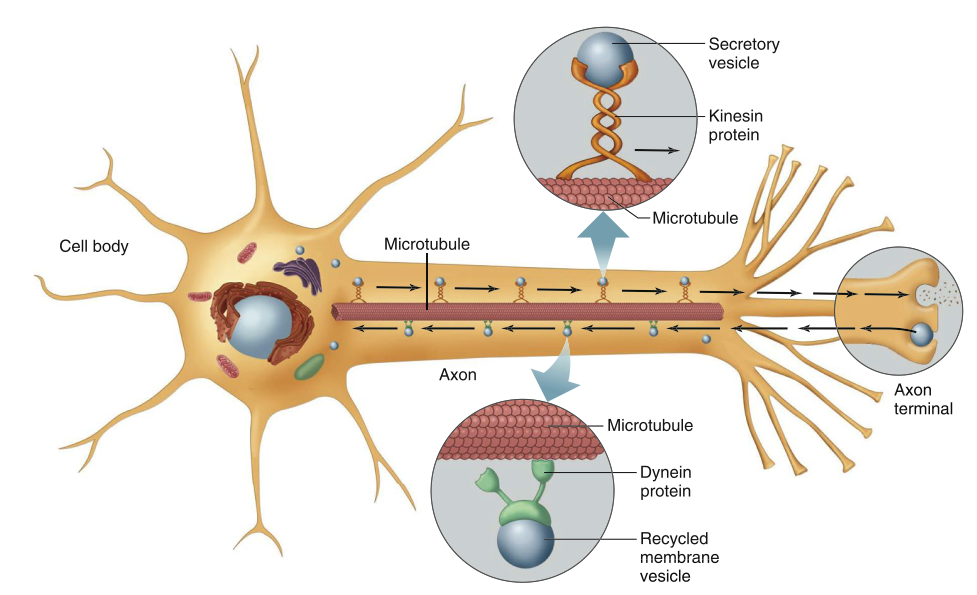
Figure 6.3. Axonal transport along microtubules by dynein and kinesin
Functional Classes of Neurons. Neurons are divided into three functional classes: afferent neurons, efferent neurons, and interneurons (Figure 6.4a). Afferent neurons convey information from the body’s tissues and organs toward the CNS. Efferent neurons transmit information away from the CNS to effector cells, including muscle, gland, or other cell types. Interneurons connect neurons within the CNS. As a rough estimate, for each afferent neuron entering the CNS, there are approximately 10 efferent neurons and 200,000 interneurons. Thus, the vast majority of neurons are interneurons.
At their peripheral ends (farthest from the CNS), afferent neurons possess sensory receptors that respond to physical or chemical changes in the environment by generating electrical signals. The receptor region may be a specialized portion of the plasma membrane or a separate cell closely associated with the neuron ending. (Recall from Chapter 5 that the term receptor has two distinct meanings: the sensory receptor defined here, and the specific proteins with which chemical messengers combine to exert effects on target cells.) Afferent neurons propagate electrical signals from their receptors into the brain or spinal cord.
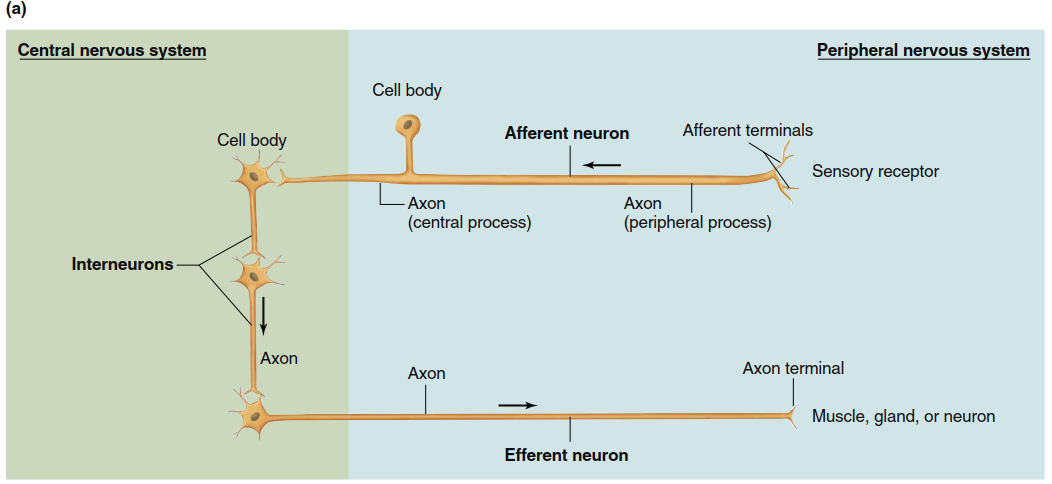
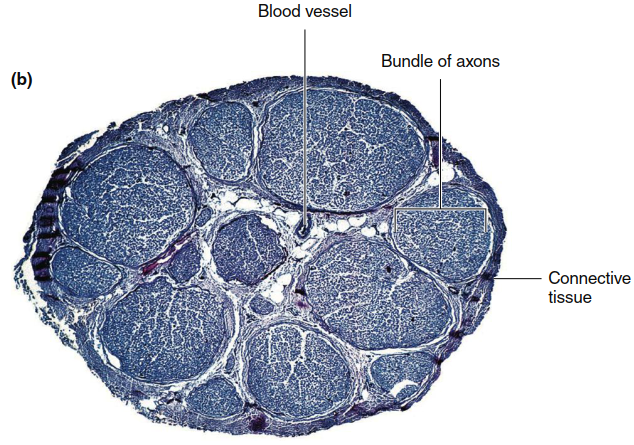
Figure 6.4. (a) Three classes of neurons. The arrows indicate the direction of transmission of neural activity. Afferent neurons in the PNS generally receive input at sensory receptors (in some cases, the afferent terminal branches themselves are modified into a sensory receptor). Efferent components of the PNS may terminate on muscle, gland, neuron, or other effector cells. Both afferent and efferent components may consist of two neurons, not one as shown here. (b) Tranverse section of a nerve as seen in a light micrograph (magnification approximately 50x). A nerve is a collection of neuron axons encased in connective tissue and is located in the peripheral nervous system
Afferent neurons have a shape distinct from that shown in Figure 6.1 because they have only a single process associated with the cell body, usually considered an axon. Shortly after leaving the cell body, the axon divides into two branches. The peripheral process begins where afferent terminal branches converge from receptor endings. The central process enters the CNS to form junctions with other neurons. As noted in Figure 6.4, for afferent neurons both the cell body and the long axon lie outside the CNS; only a portion of the central process enters the brain or spinal cord.
Efferent neurons typically exhibit the shape shown in Figure 6.1. Their cell bodies and dendrites are generally located within the CNS, while their axons extend outward to the periphery. Exceptions exist, such as in the enteric nervous system of the gastrointestinal tract (described in Chapter 15). Bundles of afferent and efferent neuron axons, along with myelin, connective tissue, and blood vessels, form the nerves of the PNS (Figure 6.4b). Note that nerve fiber refers to a single axon, whereas a nerve is a bundle of axons (fibers) bound together by connective tissue.
Interneurons lie entirely within the CNS. They account for over 99% of all neurons and exhibit a wide range of physiological properties, shapes, and functions. The number of interneurons interposed between specific afferent and efferent neurons varies according to the complexity of the action they control. For example, the knee‑jerk reflex elicited by tapping below the kneecap activates thigh muscles largely without interneurons—most afferent neurons interact directly with efferent neurons. In contrast, hearing a song or smelling a perfume that evokes memories of someone you know may involve millions of interneurons. Table 6.1 summarizes the characteristics of the three functional classes of neurons.

Synapses and Information Flow. The anatomically specialized junction between two neurons, where one neuron alters the electrical and chemical activity of another, is called a synapse. At most synapses, signals are transmitted from one neuron to another by neurotransmitters—a term that also includes the chemicals efferent neurons use to communicate with effector cells (e.g., muscle cells). Neurotransmitters released from a presynaptic neuron bind to specific protein receptors on the membrane of the receiving neuron. (Again, do not confuse this use of receptor with sensory receptors at the peripheral ends of afferent neurons.)
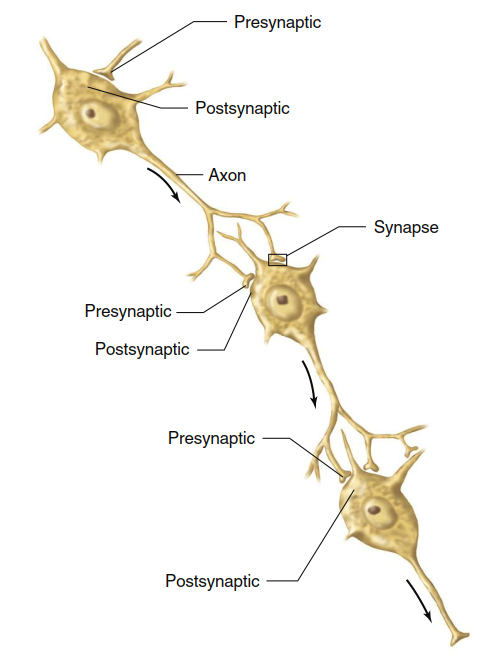
Figure 6.5. A neuron postsynaptic to one cell can be presynaptic to another. Arrows indicate direction of neural transmission
Most synapses occur between an axon terminal of one neuron and a dendrite or the cell body of a second neuron. A neuron that conducts a signal toward a synapse is termed a presynaptic neuron, whereas a neuron conducting signals away from a synapse is a postsynaptic neuron. Figure 6.5 illustrates how, in a multineuronal pathway, a single neuron can be postsynaptic to one cell and presynaptic to another. A postsynaptic neuron may have thousands of synaptic junctions on the surface of its dendrites and cell body, allowing signals from many presynaptic neurons to affect it. Interconnected in this way, the millions of neurons in the nervous system exemplify the general principle of physiology that information flow between cells, tissues, and organs is essential for homeostasis and enables complex integration of physiological processes.
Date added: 2026-07-14; views: 3;