Neuron Electrical Signaling: Resting Membrane Potential and Ohm’s Law in Physiology
This section provides an excellent demonstration of the general principle that physiological processes are dictated by the laws of chemistry and physics, notably those governing the net flux of charged molecules. As discussed in Chapter 4, the predominant solutes in the extracellular fluid are sodium ions (Na⁺) and chloride ions (Cl⁻). In contrast, the intracellular fluid contains high concentrations of potassium ions (K⁺) along with ionized nonpenetrating molecules, particularly phosphate compounds and proteins with negatively charged side chains. Electrical phenomena resulting from the distribution of these charged particles occur at the cell’s plasma membrane and play a critical role in signal integration and cell-to-cell communication—the two major functions of neurons.
A fundamental physical principle states that like charges repel each other: positive repels positive, and negative repels negative. Conversely, oppositely charged substances attract and will move toward each other unless separated by a barrier (Figure 6.7). Separated electrical charges of opposite sign possess the potential to do work if allowed to come together. This potential is called an electrical potential or, because it is determined by the difference in charge between two points, a potential difference (often referred to simply as the potential). The unit of electrical potential is the volt. In most biological systems, the total charge that can be separated is very small, so potential differences are measured in millivolts (1 mV = 0.001 V).
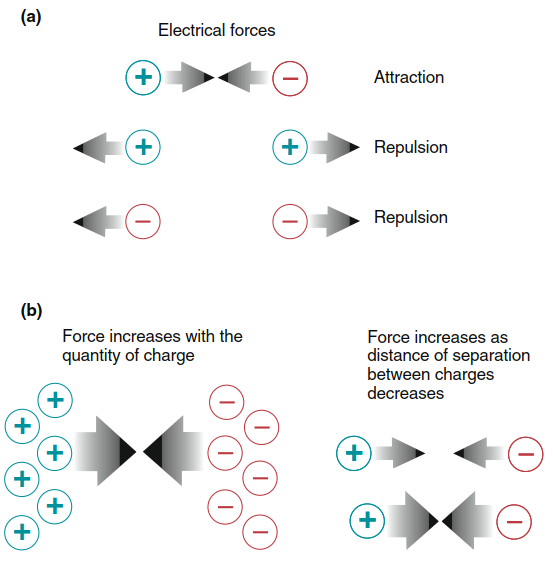
Figure 6.7. (a) Types of electrical interactions. (b) Effects on electrical forces of quantity and distance between charges
The movement of electrical charge is termed a current. The electrical potential between charges tends to make them flow, producing a current. Opposite charges produce a current that brings them toward each other; like charges produce a current that increases separation. The magnitude of the current depends on the potential difference between the charges and on the resistance of the material or structure through which they move. Resistance is the hindrance to electrical charge movement—high resistance yields low current flow. The relationship between voltage (V), resistance (R), and current (I) is expressed by Ohm’s law: I = V / R. Materials with high electrical resistance are called insulators, while those with low resistance are conductors.
Water containing dissolved ions is a relatively good conductor because ions can carry the current. As noted, intracellular and extracellular fluids contain many ions and thus conduct current readily. Lipids, however, contain very few charged groups and cannot carry current. Therefore, the lipid layers of the plasma membrane are regions of high electrical resistance that separate the intracellular fluid from the extracellular fluid—two low-resistance aqueous compartments. This arrangement is fundamental to the generation of the resting membrane potential, typically ranging from –40 mV to –90 mV in different neurons.
The Resting Membrane Potential: Origin and Maintenance. The resting membrane potential arises from the unequal distribution of ions across the plasma membrane and the membrane’s selective permeability. At rest, the neuronal membrane is much more permeable to K⁺ than to Na⁺ due to the presence of leak potassium channels. Because the intracellular fluid contains a high concentration of K⁺ and the extracellular fluid contains a low concentration of K⁺, potassium ions diffuse down their concentration gradient out of the cell. This movement of positive charge outward creates a negative charge inside relative to the outside, establishing the resting potential.
The sodium‑potassium pump (Na⁺/K⁺‑ATPase) actively maintains these ion gradients by transporting three Na⁺ out of the cell and two K⁺ into the cell for each molecule of ATP hydrolyzed. This electrogenic pump contributes directly (about –5 to –10 mV) to the resting potential and indirectly by maintaining the concentration gradients that drive passive ion fluxes. Without this pump, the gradients would gradually dissipate, and the neuron would lose its ability to generate electrical signals.
Electrochemical Gradients and Driving Forces. The net movement of an ion across the membrane depends on both its concentration gradient (chemical force) and the membrane potential (electrical force). The combined influence is called the electrochemical gradient. For each ion, the equilibrium potential—calculated using the Nernst equation—is the membrane potential at which the electrical force exactly opposes the chemical force, resulting in no net ion movement. At rest, the membrane potential is close to the equilibrium potential for K⁺ (about –90 mV) but far from that for Na⁺ (about +60 mV), reflecting the membrane’s high resting permeability to K⁺.
Local Potentials, Action Potentials, and Signal Propagation. When a neuron receives a synaptic input, neurotransmitters open ligand‑gated ion channels, causing brief local changes in membrane potential called graded potentials (or local potentials). If the membrane is depolarized to a critical threshold (approximately –55 mV), voltage‑gated sodium channels open rapidly, allowing an influx of Na⁺ that generates a rapid, all‑or‑none action potential. The action potential propagates along the axon without decrement, driven by the sequential opening of voltage‑gated Na⁺ channels followed by voltage‑gated K⁺ channels that repolarize the membrane.
Ohm’s law (I = V/R) governs the passive spread of electrical signals within neurons. The length constant (λ) describes how far a graded potential can travel along a dendrite or axon before decaying; it is directly proportional to the square root of membrane resistance and inversely proportional to the square root of internal resistance. Myelination increases membrane resistance and decreases capacitance, thereby increasing the length constant and conduction velocity—a principle illustrated by the faster conduction in myelinated axons where action potentials jump between nodes of Ranvier (saltatory conduction).
Clinical Relevance and Experimental Approaches. Disruptions in ion channel function, known as channelopathies, can cause neurological disorders such as epilepsy, migraine, and periodic paralysis. Local anesthetics (e.g., lidocaine) block voltage‑gated sodium channels, preventing action potential propagation and thereby eliminating pain signals. Understanding the electrical properties of neurons also underpins electrophysiological techniques like patch‑clamp recording, which allows direct measurement of ion currents through single channels, and electroencephalography (EEG) , which records summed postsynaptic potentials from large populations of cortical neurons.
In summary, the physical principles of charge interaction, potential difference, resistance, and current flow—encapsulated in Ohm’s law—are directly applicable to neuronal signaling. The plasma membrane’s lipid bilayer acts as an insulator separating two conductive aqueous compartments, enabling the rapid, controlled movement of ions that underlies all nervous system function, from sensory transduction to complex cognition.
Date added: 2026-07-14; views: 3;