Resting Membrane Potential, Graded Potentials, and Action Potentials: A Comprehensive Guide to Excitable Membranes
All cells possess a resting membrane potential due to the presence of ion pumps, ion concentration gradients, and leak channels within the cell membrane. However, certain cells contain an additional group of ion channels that can be gated (opened or closed) under specific conditions, enabling the generation of electrical signals for information transmission across different membrane regions. This property is termed excitability, and such membranes are referred to as excitable membranes. Neurons and muscle cells represent the primary cell types exhibiting this characteristic. Electrical signals manifest in two distinct forms: graded potentials and action potentials, where graded potentials facilitate short-distance signaling, while action potentials enable long-distance communication, particularly crucial in neuronal and muscle cell membranes.
Directional Changes in Membrane Potential: Depolarization, Repolarization, and Hyperpolarization. The terms depolarize, repolarize, and hyperpolarize describe directional changes in membrane potential relative to the resting potential in excitable cells (Figure 6.14). The resting membrane potential is described as polarized, indicating a net charge difference between the cell's exterior and interior. Depolarization occurs when the membrane potential becomes less negative (closer to zero) than the resting level, while overshoot refers to a reversal of polarity where the cell interior becomes positive relative to the exterior. Repolarization describes the return of a depolarized membrane to its resting value, and hyperpolarization occurs when the potential becomes more negative than the resting level.
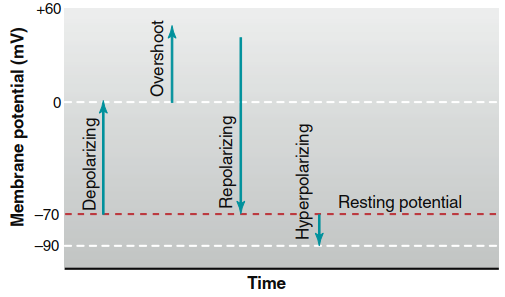
Figure 6.14. Depolarizing, repolarizing, hyperpolarizing, and overshoot changes in membrane potential relative to the resting potential
Ionic Mechanisms Underlying Membrane Potential Changes. Changes in membrane potential that neurons utilize as signals arise from alterations in the cell membrane's permeability to ions. As introduced in Chapter 4, gated ion channels may be opened or closed by mechanical, electrical, or chemical stimuli. When a neuron receives a chemical signal from an adjacent neuron, certain gated channels open, permitting greater ionic current across the membrane. The increased movement of ions down their electrochemical gradient modifies the membrane potential, resulting in either depolarization or hyperpolarization relative to the resting state. The specific characteristics of these gated ion channels ultimately determine the nature of the generated electrical signal.
Graded Potentials: Localized Membrane Potential Changes. Graded potentials represent changes in membrane potential confined to a relatively small region of the plasma membrane, typically produced when specific environmental changes act on specialized membrane regions. They receive their name because the magnitude of the potential change can vary (is "graded"), and they are classified by location or function—including receptor potentials, synaptic potentials, and pacemaker potentials (Table 6.3).

Whenever a graded potential occurs, charge flows between the origin site and adjacent membrane regions still at resting potential. In Figure 6.15, transient application of a chemical signal depolarizes a small membrane region by briefly opening membrane cation channels, producing a less negative potential than neighboring areas. Positive intracellular charges (primarily K⁺ ions) move through the intracellular fluid away from the depolarized region toward more negative, resting membrane areas, while simultaneously, extracellular positive charge moves from more positive resting regions toward less positive depolarized areas.
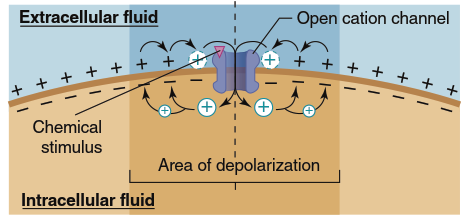
Figure 6.15. Depolarization and graded potential caused by a chemical stimulus. Inward positive current through ligand-gated cation channels depolarizes a region of the membrane, and local currents spread the depolarization to adjacent regions
Depending on the initiating event, graded potentials can occur in either depolarizing or hyperpolarizing directions (Figure 6.16a), with their magnitude related to the magnitude of the initiating event (Figure 6.16b). In addition to ion movement inside and outside the cell, charge is lost across the membrane due to permeability through open leak channels. Consequently, the change in membrane potential decreases as distance from the initial potential change site increases (Figure 6.16c). Plasma membranes are so leaky to ions that these currents die out almost completely within a few millimeters of their origin, making local current decremental—charge flow decreases with increasing distance from the graded potential origin (Figure 6.17).
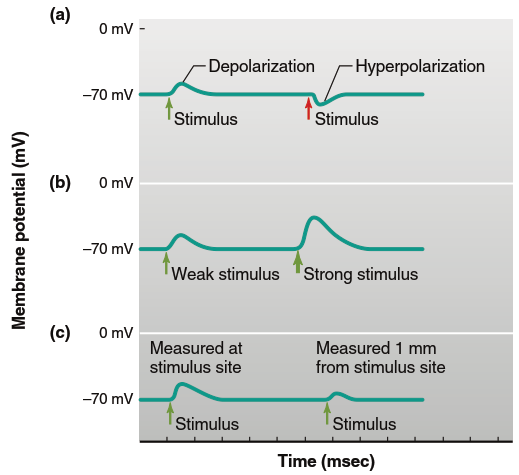
Figure 6.16. Graded potentials can be recorded under experimental conditions in which the stimulus strength can vary. Such experiments show that graded potentials (a) can be depolarizing or hyperpolarizing, (b) can vary in size, and (c) are conducted decrementally. In this example, the resting membrane potential is 270 mV
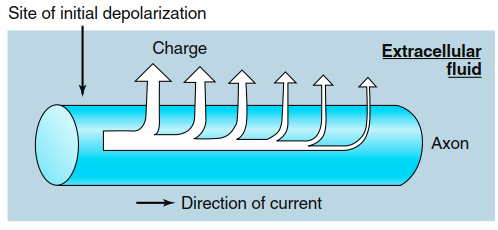
Figure 6.17. Leakage of charge (predominately K+) across the plasma membrane reduces the local current at sites farther along the membrane from the site of initial depolarization
Because the electrical signal diminishes with distance, graded potentials and their associated local current can function as signals only over very short distances (a few millimeters). However, if additional stimuli occur before the graded potential has dissipated, they can add to the depolarization from the first stimulus through a process called summation, which is particularly important for sensation (discussed in Chapter 7). Graded potentials serve as the only communication means for some neurons, while in other neurons, they initiate longer-distance signals described next.
Action Potentials: Large-Scale, Rapid Membrane Potential Changes. Action potentials differ substantially from graded potentials, representing large alterations in membrane potential that may change by as much as 100 mV—for example, depolarizing from -70 to +30 mV and then repolarizing to resting potential. These signals are generally very rapid (as brief as 1-4 milliseconds) and may repeat at frequencies of several hundred per second. The propagation of action potentials down the axon constitutes the mechanism by which the nervous system communicates from cell to cell over long distances. Understanding the ion channel properties that enable these large, rapid changes and how action potentials propagate along excitable membranes requires examination of voltage-gated channels.
Voltage-Gated Ion Channels: Structure and Function. As introduced in Chapter 4, multiple ion channel types exist with different mechanisms regulating their opening. Ligand-gated ion channels open in response to signaling molecule binding (as shown in Figure 6.15), while mechanically gated ion channels open following physical deformation (stretching) of the plasma membrane. Although these channel types often mediate graded potentials that can serve as initiating stimuli for action potentials, it is voltage-gated ion channels that confer upon membranes the ability to undergo action potentials. Dozens of voltage-gated ion channel types exist, varying by conducted ion (e.g., Na⁺, K⁺, Ca²⁺, or Cl⁻) and behavioral characteristics as membrane voltage changes. This discussion focuses on the specific voltage-gated Na⁺ and K⁺ channels that mediate most neuronal action potentials.
Figure 6.18 summarizes the relevant characteristics of these channels. Both Na⁺ and K⁺ channels contain sequences of charged amino acid residues in their structure, enabling reversible shape changes in response to membrane potential alterations. At negative membrane potentials (e.g., at resting membrane potential), both channel types tend to close, while membrane depolarization tends to open them. Two key differences allow these channels to contribute differently to action potential production. First, voltage-gated Na⁺ channels respond faster to changes in membrane voltage—when a membrane area suddenly depolarizes, local voltage-gated Na⁺ channels open before voltage-gated K⁺ channels, and if the membrane repolarizes to negative voltages, voltage-gated K⁺ channels are slower to close. Second, voltage-gated Na⁺ channels possess an extra structural feature called an inactivation gate, visualized as a "ball and chain," which limits Na⁺ flux by blocking the channel shortly after depolarization opens it. When the membrane repolarizes, the channel closes, forcing the inactivation gate back out of the pore and allowing channel return to the closed state.
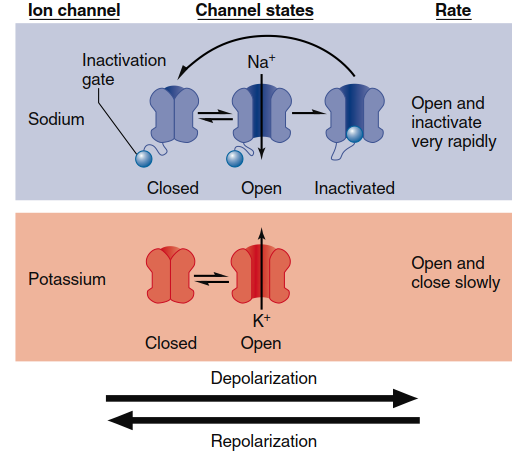
Figure 6.18. Behavior of voltage-gated Na+ and K+ channels. Depolarization of the membrane causes Na+ channels to rapidly open, then undergo inactivation followed by the opening of K+ channels. When the membrane repolarizes to negative voltages, both channels return to the closed state
Action Potential Mechanism: Step-by-Step Process. Integrating these channel properties with basic principles governing membrane potentials explains how action potentials occur. During an action potential, transient changes in membrane permeability allow Na⁺ and K⁺ to move down their electrochemical gradients. Figure 6.19 illustrates the sequential steps. In step 1, the resting membrane potential is close to the K⁺ equilibrium potential because more open K⁺ channels (leak channels, distinct from voltage-gated channels) exist than Na⁺ channels. An action potential begins with a depolarizing stimulus—for example, when a neurotransmitter binds to a specific ligand-gated ion channel, allowing Na⁺ to enter the cell (review Figure 6.15). This initial depolarization stimulates the opening of some voltage-gated Na⁺ channels, and further Na⁺ entry through those channels adds to local membrane depolarization.
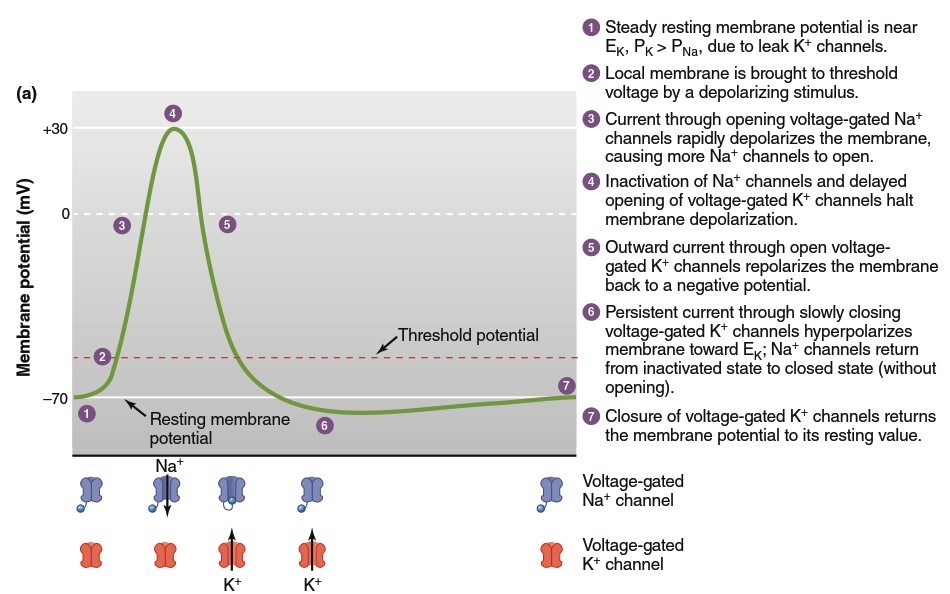
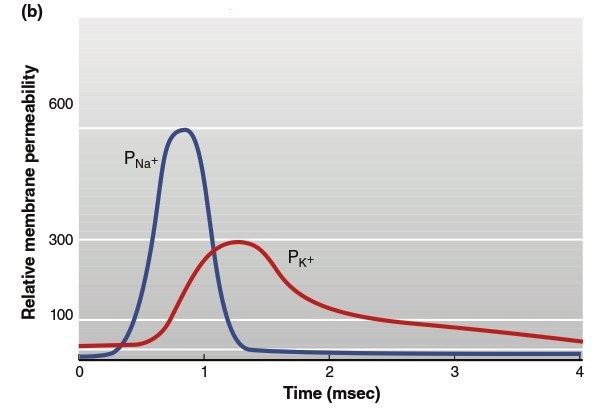
Figure 6.19. The changes in (a) membrane potential and (b) relative membrane permeability (P) to sodium and potassium ions during an action potential. Steps 1–7 are described in more detail in the text
When the membrane reaches a critical threshold potential (step 2), depolarization becomes a positive feedback loop: Na⁺ entry causes depolarization, which opens more voltage-gated Na⁺ channels, causing more depolarization, and so on. This process produces rapid membrane potential depolarization (step 3), with overshoot making the interior positive and exterior negative. In this phase, the membrane approaches but does not quite reach the Na⁺ equilibrium potential (+60 mV). As the membrane potential reaches its peak value (step 4), Na⁺ permeability abruptly declines as inactivation gates break the positive feedback cycle by blocking open Na⁺ channels. Simultaneously, the depolarized membrane state opens relatively sluggish voltage-gated K⁺ channels, and the resulting elevated K⁺ flux out of the cell rapidly repolarizes the membrane toward its resting value (step 5).
The return to negative membrane potential causes voltage-gated Na⁺ channels to transition from their inactivated state back to the closed state (without opening), and K⁺ channels to also return to closed state. Because voltage-gated K⁺ channels close relatively slowly, immediately after an action potential there is a period when K⁺ permeability remains above resting levels, and the membrane transiently hyperpolarizes toward the K⁺ equilibrium potential (step 6)—this portion is known as the afterhyperpolarization. Once voltage-gated K⁺ channels finally close, the resting membrane potential is restored (step 7). Whereas voltage-gated Na⁺ channels operate in positive feedback mode at the action potential's beginning, voltage-gated K⁺ channels terminate the action potential and induce their own closing through a negative feedback process (Figure 6.20).
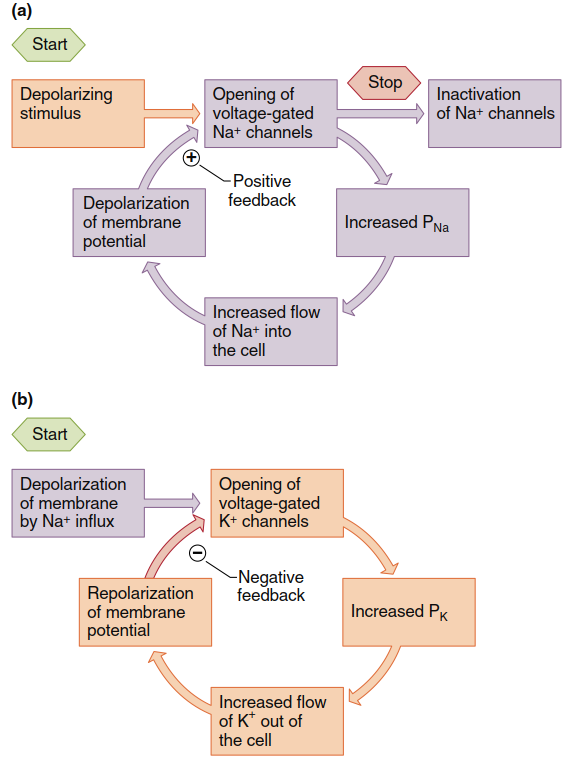
Figure 6.20. Feedback control in voltage-gated ion channels. (a) Na+ channels exert positive feedback on membrane potential. (b) K+ channels exert negative feedback
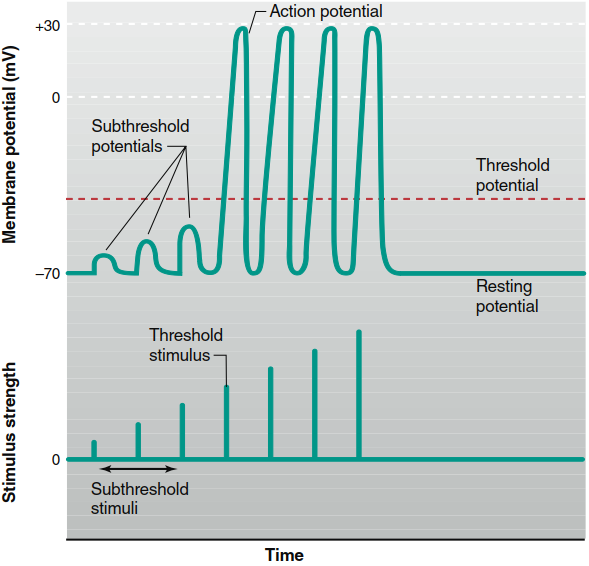
Figure 6.21. Changes in the membrane potential with increasing strength of excitatory stimuli. When the membrane potential reaches threshold, action potentials are generated. Increasing the stimulus strength above threshold level does not cause larger action potentials. (The absolute value of threshold is not indicated because it varies from cell to cell.)
Ion Flux and Concentration Gradient Maintenance. Despite the large changes in membrane potential, the number of ions crossing the membrane during a single action potential is extremely small compared to total cellular ions, producing only infinitesimal changes in intracellular ion concentrations. However, without eventual movement back across the membrane, the concentration gradients of Na⁺ and K⁺ would gradually dissipate with repeated action potentials, preventing further action potential generation. Cellular accumulation of Na⁺ and loss of K⁺ are prevented by the continuous action of membrane Na⁺/K⁺-ATPase pumps.
Threshold, All-or-None Principle, and Stimulus Strength. Not all membrane depolarizations in excitable cells trigger the positive feedback process leading to an action potential. Action potentials occur only when the initial stimulus plus current through opened Na⁺ channels sufficiently elevate the membrane potential beyond the threshold potential. Stimuli just strong enough to depolarize the membrane to this level are threshold stimuli (Figure 6.21). The threshold of most excitable membranes is approximately 15 mV less negative than the resting membrane potential—for a neuron with -70 mV resting potential, threshold potential may be -55 mV. At depolarizations below threshold, the positive feedback cycle cannot initiate, and the membrane returns to resting level upon stimulus removal without generating an action potential; these weak depolarizations are called subthreshold potentials, caused by subthreshold stimuli.
Stimuli stronger than those required to reach threshold elicit action potentials, but as shown in Figure 6.21, the resulting action potentials have exactly the same amplitude as those caused by threshold stimuli. Once threshold is reached, membrane events no longer depend upon stimulus strength because depolarization generates an action potential through the operating positive feedback cycle. Action potentials either occur maximally or not at all—a principle described as all-or-none. The firing of a gun provides a mechanical analogy: the explosion magnitude and bullet velocity do not depend on how hard the trigger is squeezed; either the trigger is pulled hard enough to fire the gun, or it is not. Because a single action potential's amplitude does not vary proportionally with stimulus amplitude, action potentials cannot convey information about initiating stimulus magnitude. Instead, as discussed later, this information depends upon the number and patterns of action potentials transmitted per unit time (frequency), not their magnitude.
Pharmacological and Toxicological Modulation. Local anesthetics such as procaine (Novocaine) and lidocaine (Xylocaine) prevent action potential generation by blocking voltage-gated Na⁺ channels, preventing their opening in response to depolarization. Without action potentials, graded signals generated in sensory neurons—for example, in response to injury—cannot reach the brain to produce pain sensation. Some animals produce toxins that interfere with nerve conduction similarly. For instance, certain organs of the pufferfish produce an extremely potent toxin, tetrodotoxin, which binds to voltage-gated Na⁺ channels and prevents the Na⁺ component of action potentials. In Japan, chefs preparing the pufferfish dish called fugu are specially trained to completely remove toxic organs before serving; individuals eating improperly prepared fugu may die even from tiny quantities of tetrodotoxin.
Refractory Periods: Absolute and Relative. During an action potential, a second stimulus—no matter how strong—will not produce a second action potential (Figure 6.22), placing that membrane region in its absolute refractory period. This occurs when voltage-gated Na⁺ channels are either already open or have progressed to the inactivated state during the first action potential. The inactivation gate blocking these channels must be removed by repolarizing the membrane and closing the pore before channels can reopen to a second stimulus. Following the absolute refractory period, an interval exists during which a second action potential can be produced—but only if stimulus strength is considerably greater than usual. This is the relative refractory period, lasting up to 15 msec and coinciding roughly with the afterhyperpolarization period.
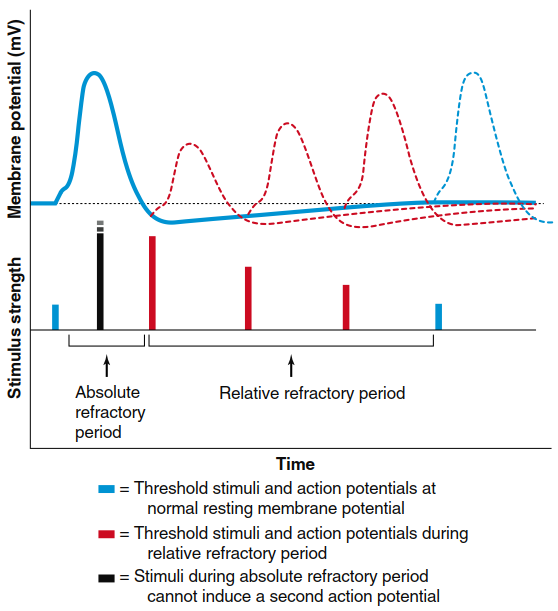
Figure 6.22. Absolute and relative refractory periods of the action potential determined by a paired-pulse protocol. After a threshold stimulus that results in an action potential (first stimulus and solid voltage trace), a second stimulus given at various times after the first can be used to determine refractory periods. All stimuli shown are of the minimum size needed to stimulate an action potential. During the absolute refractory period, a second stimulus (black), no matter how strong, will not produce a second action potential. In the relative refractory period (stimuli and action potentials shown in red), a second action potential can be triggered, but a larger stimulus is required to reach threshold, mainly because K+ permeability is still above resting levels. Action potentials are reduced in size during the relative refractory period, due both to the residual inactivation of some Na+ channels and the persistence of some open K+ channels.
During the relative refractory period, some but not all voltage-gated Na⁺ channels have returned to a resting state; with fewer Na⁺ channels available, action potential magnitude is temporarily reduced. Additionally, some K⁺ channels that repolarized the membrane remain open. Outflow of K⁺ through these channels opposes some depolarization produced by Na⁺ entry, making threshold more difficult to reach unless a stronger stimulus occurs. Thus, during the relative refractory state, a new stimulus can depolarize the membrane above threshold potential only if the stimulus is large in magnitude or outlasts the relative refractory period. Refractory periods limit the number of action potentials an excitable membrane can produce per unit time. Most neurons respond at frequencies up to 100 action potentials per second, with some producing higher frequencies briefly. Refractory periods contribute to action potential separation so individual electrical signals pass down the axon and are key in determining propagation direction.
Action Potential Propagation Mechanisms. The action potential can travel the length of a neuron only if each membrane point is depolarized to threshold as the action potential moves down the axon (Figure 6.23). As with graded potentials (refer back to Figure 6.15), the membrane is depolarized at each point relative to adjacent membrane portions still at resting potential. The potential difference causes local current to flow, depolarizing adjacent membrane where it opens voltage-gated Na⁺ channels. The current entering during an action potential sufficiently depolarizes adjacent membrane to threshold potential. The new action potential produces its own local currents depolarizing the next adjacent region (Figure 6.23b), generating another action potential at the subsequent site, thereby causing sequential opening and closing of voltage-gated Na⁺ and K⁺ channels along the membrane.
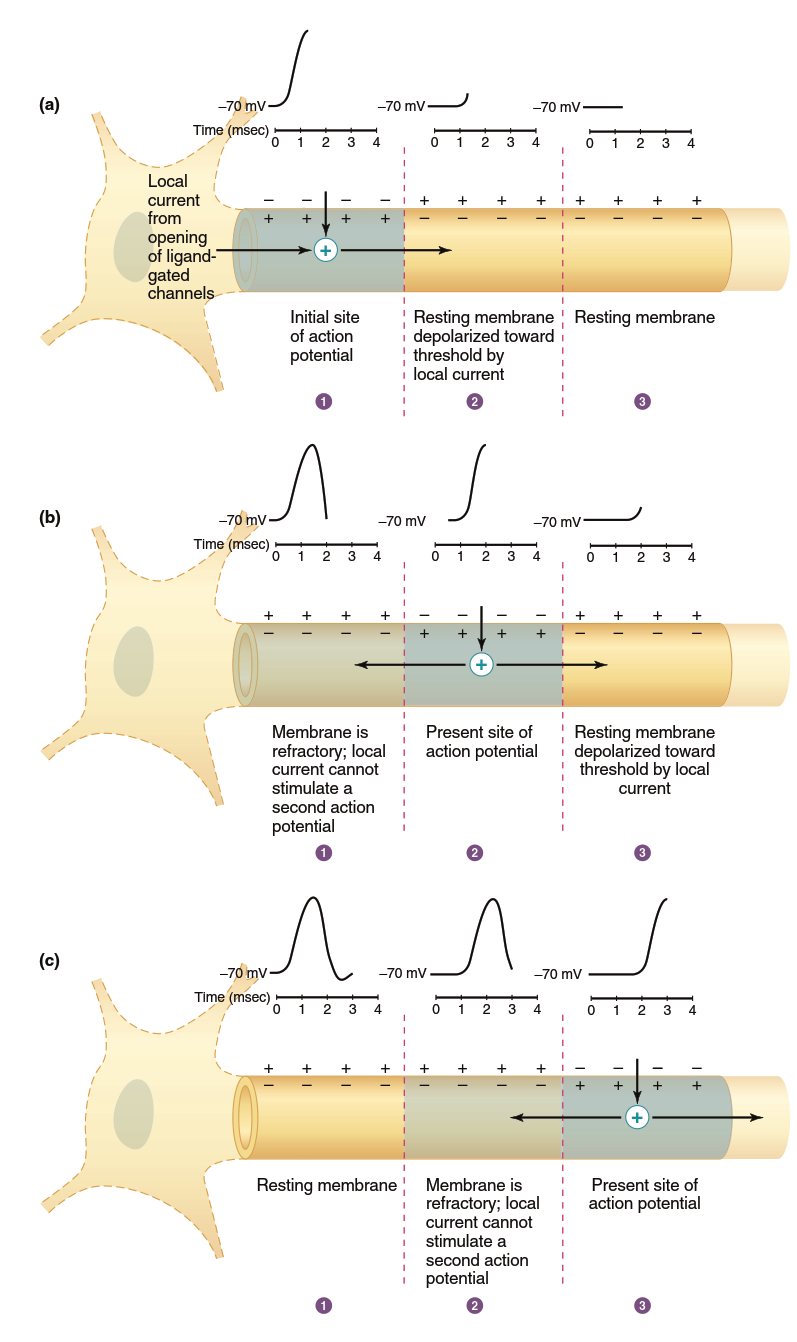
Figure 6.23. One-way propagation of an action potential. For simplicity, potentials are shown only on the upper membrane, local currents are shown only on the inside of the membrane, and repolarizing currents are not shown. (a) Local current from the opening of ligand-gated ion channels in the cell body and dendrites causes an action potential to be initiated in region 1, and local current depolarizes region 2. (b) Action potential in region 2 generates local currents; region 3 is depolarized toward threshold, but region 1 is refractory. (c) Action potential in region 3 generates local currents, but region 2 is refractory
This process resembles lighting a trail of gunpowder—the action potential does not physically move but "sets off" a new action potential in the axon region just ahead. Because each action potential regeneration depends on the positive feedback cycle of a new group of Na⁺ channels where the action potential occurs, the action potential arriving at the membrane end is virtually identical in form to the initial one. Therefore, action potentials are not decremental; they do not decrease in magnitude with distance like graded potentials. Because a membrane area that has just undergone an action potential is refractory and cannot immediately undergo another, the only direction of propagation is away from recently active membrane regions—again similar to burning gunpowder, where fire spreads only forward where gunpowder is fresh, not backward where it has already burned.
If the membrane through which the action potential must travel is not refractory, excitable membranes can conduct action potentials in either direction, with propagation direction determined by stimulus location. For example, action potentials in skeletal muscle cells initiate near the middle of cells and propagate toward both ends. In most neurons, however, action potentials initiate at one cell end and propagate toward the other end, as shown in Figure 6.23. Propagation ceases when the action potential reaches an axon end.
Factors Affecting Conduction Velocity: Diameter and Myelination. The velocity of action potential propagation along a membrane depends upon fiber diameter and whether the fiber is myelinated. Larger fiber diameter yields faster propagation because a wide fiber offers less internal resistance to local current; more ions flow in a given time, bringing adjacent membrane regions to threshold faster. Myelin acts as an insulator, making charge flow between intracellular and extracellular fluid compartments more difficult. Because less charge "leaks" across myelin, local current can spread farther along an axon. Moreover, the concentration of voltage-gated Na⁺ channels in myelinated axon regions is low; therefore, action potentials occur only at nodes of Ranvier, where the myelin coating is interrupted and voltage-gated Na⁺ channel concentration is high (Figure 6.24).
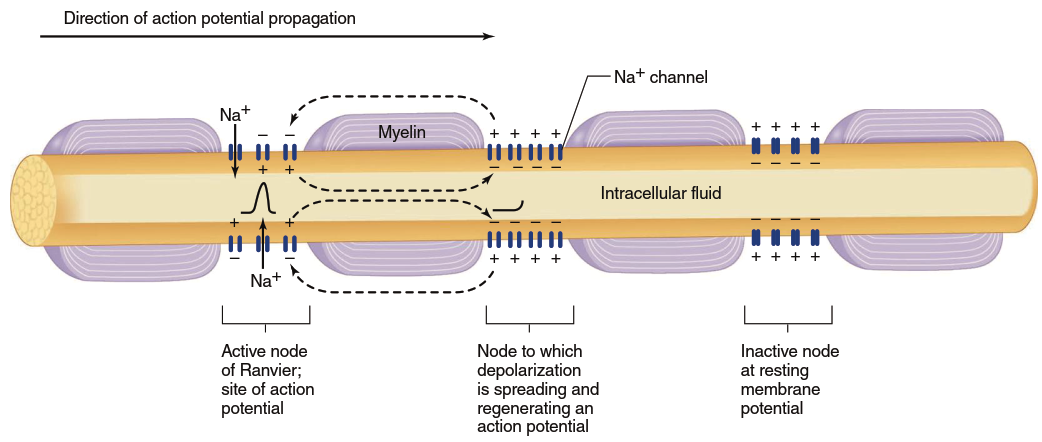
Figure 6.24. Myelinization and saltatory conduction of action potentials. K+ channels are not depicted (they are located primarily at the myelin/node junctions and help to repolarize the neuron)
Action potentials appear to "jump" from one node to the next as they propagate along a myelinated fiber, leading to the term saltatory conduction (Latin saltare, "to leap"). However, it is important to understand that an action potential does not actually jump but rather is regenerated at each node. Propagation via saltatory conduction is faster than in nonmyelinated fibers of the same axon diameter because less charge leaks out through myelin-covered membrane sections, more charge arrives at the node adjacent to the active node, and an action potential generates there sooner than without myelin. Furthermore, because ions cross the membrane primarily at nodes of Ranvier, membrane pumps need to restore fewer ions. Myelinated axons are metabolically more efficient than unmyelinated ones, adding speed, reducing metabolic cost, and saving nervous system space because axons can be thinner.
Conduction velocities range from approximately 0.5 m/sec (1 mi/h) for small-diameter, unmyelinated fibers to about 100 m/sec (225 mi/h) for large-diameter, myelinated fibers. At 0.5 m/sec, an action potential would travel from the toe to the spinal cord and brain of an average-sized person in about 4 seconds; at 100 m/sec, it takes only about 0.02 seconds. This explains why dropping a heavy object on your toe produces an immediate, sharp pain (carried by large-diameter, myelinated neurons) before the onset of a dull, throbbing ache (transmitted along small-diameter, unmyelinated neurons).
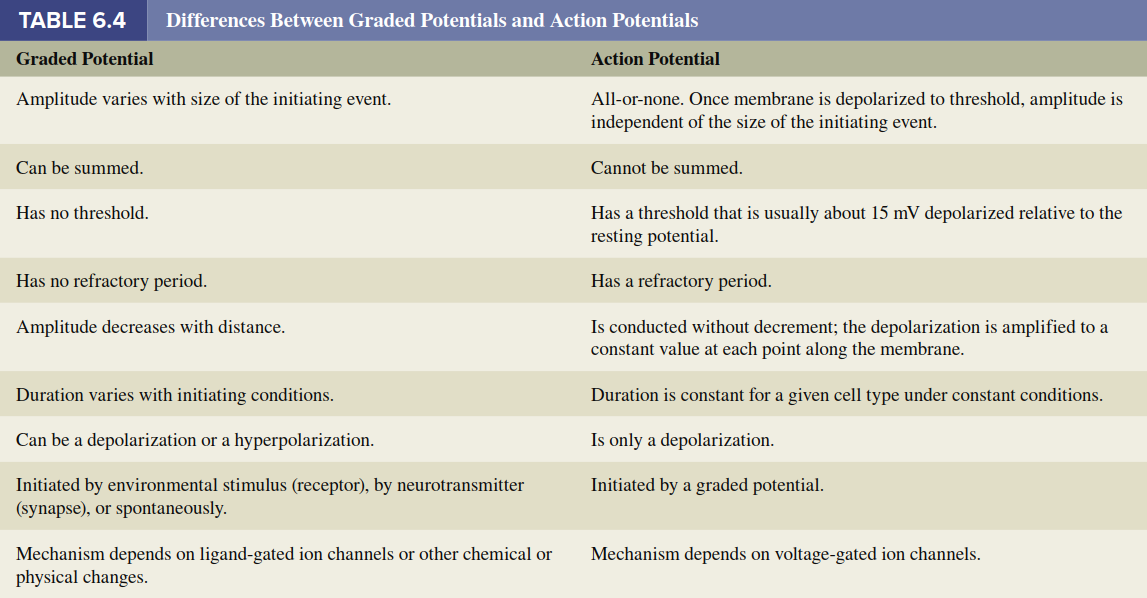
Generation of Action Potentials in Different Neuron Types. In our description of action potentials thus far, "stimuli" have been discussed as initiators bringing the membrane to threshold potential, where voltage-gated Na⁺ channels initiate the action potential. In afferent neurons, the initial depolarization to threshold is achieved by a graded potential called a receptor potential, generated in sensory receptors at the peripheral ends of neurons (the ends farthest from the CNS). In all other neurons, depolarization to threshold occurs either due to a graded potential generated by synaptic input to the neuron (known as a synaptic potential) or to a spontaneous change in membrane potential (known as a pacemaker potential). The next section addresses synaptic potential production; Chapter 7 discusses receptor potential production; and Chapters 12, 13, and 15 consider pacemaker potentials in different organ systems. The differences between graded potentials and action potentials are summarized in Table 6.4.
Date added: 2026-07-14; views: 3;