Synaptic Transmission: Neurotransmitter Release, Receptor Binding, and Postsynaptic Potentials
As illustrated in Figure 6.27a, neurotransmitters are stored within small vesicles enclosed by lipid bilayer membranes. Prior to neuronal activation, many vesicles are docked at specialized release sites called active zones on the presynaptic membrane, while others remain dispersed within the terminal. Neurotransmitter release is initiated when an action potential reaches the presynaptic terminal membrane. A key feature of chemical synapse terminals is the presence of voltage-gated Ca²⁺ channels in addition to the Na⁺ and K⁺ channels found elsewhere in the neuron. Depolarization during the action potential opens these Ca²⁺ channels, and because the electrochemical gradient favors Ca²⁺ influx, calcium ions flow into the axon terminal.
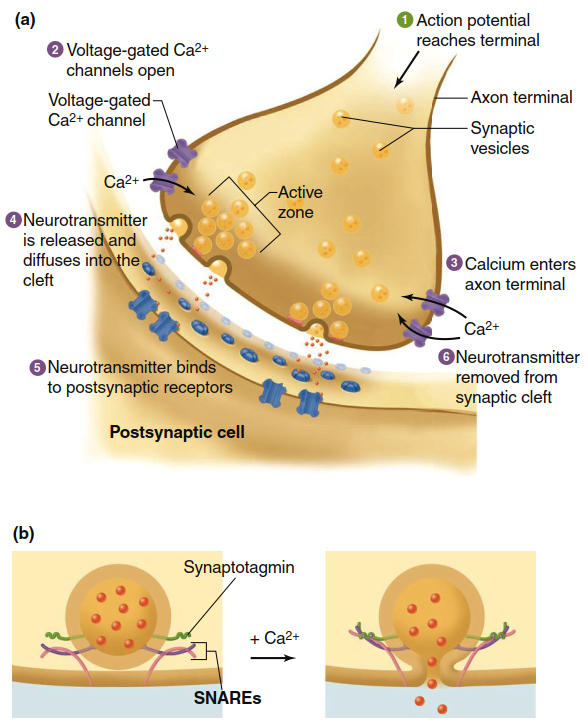
Figure 6.27. (a) Mechanisms of signaling at a chemical synapse. (b) Magnified view showing details of neurotransmitter release. Calcium ions trigger synaptotagmin and SNARE proteins to induce membrane fusion and neurotransmitter release. (SNARE 5 Soluble N-ethylmaleimide-sensitive factor attachment protein receptor)
Calcium ions activate processes that lead to the fusion of docked vesicles with the synaptic terminal membrane (Figure 6.27b). Before action potential arrival, vesicles are loosely docked in active zones through interactions among SNARE proteins (soluble N-ethylmaleimide-sensitive factor attachment protein receptors), some anchored in the vesicle membrane and others in the terminal membrane. Calcium ions entering during depolarization bind to synaptotagmins, a family of vesicle-associated proteins, triggering a conformational change in the SNARE complex that causes membrane fusion and neurotransmitter release. After fusion, vesicles may undergo two possible fates: at some synapses, they completely fuse and are later recycled via endocytosis from membrane sites outside the active zone (see Figure 4.21). At other synapses, especially those with high-frequency action potential firing, vesicles may briefly fuse, release their contents, reseal the pore, and withdraw into the axon terminal through a mechanism called “kiss-and-run fusion.”
Once released from the presynaptic axon terminal, neurotransmitters diffuse across the synaptic cleft to interact with the postsynaptic cell. A fraction of these neurotransmitter molecules bind to receptors on the postsynaptic plasma membrane. Activated receptors may be ionotropic receptors (directly functioning as ion channels, as reviewed in Figure 6.15) or metabotropic receptors (indirectly influencing ion channels via G proteins and/or second messengers). In either case, neurotransmitter binding results in the opening or closing of specific ligand-gated ion channels in the postsynaptic membrane, ultimately producing changes in the membrane potential. Due to the sequence of events, a brief synaptic delay of approximately 0.2 milliseconds occurs between action potential arrival at the presynaptic terminal and the resulting membrane potential changes in the postsynaptic cell.
Neurotransmitter binding to receptors is transient and reversible, existing in equilibrium between bound and unbound forms. As unbound neurotransmitter concentration in the synaptic cleft decreases, the number of occupied receptors correspondingly declines. When neurotransmitters are no longer bound, ion channels in the postsynaptic membrane return to their resting state. This reversibility ensures precise temporal control of synaptic signaling.
Removal of Neurotransmitter from the Synapse. Presynaptic cells typically secrete large amounts of neurotransmitter to maximize the likelihood of binding to postsynaptic receptors. However, unbound neurotransmitters must be removed to terminate the signal and prevent diffusion to nearby cells where they might cause unintended effects. Unbound neurotransmitters are cleared from the synaptic cleft through four primary mechanisms: (1) active transport back into the presynaptic axon terminal for reuse (reuptake); (2) transport into nearby glial cells for degradation; (3) diffusion away from the receptor site; or (4) enzymatic transformation into inactive substances, some of which are transported back into the presynaptic terminal for reuse. The enzymes involved in enzymatic transformation may be located on the postsynaptic membrane, presynaptic membrane, or within the synaptic cleft.
Excitatory Chemical Synapses. The two types of chemical synapses—excitatory and inhibitory—are distinguished by their effect on the postsynaptic cell, which depends on the ion channels influenced by neurotransmitter binding. At an excitatory chemical synapse, the postsynaptic response is a depolarization that brings the membrane potential closer to threshold. The usual effect of activated receptors at such synapses is to open nonselective channels permeable to both Na⁺ and K⁺. Electrical and concentration gradients drive Na⁺ into the cell, while for K⁺, the electrical gradient opposes the concentration gradient (review Figure 6.12).
Opening channels permeable to both ions results in simultaneous movement of a relatively small number of K⁺ ions out of the cell and a larger number of Na⁺ ions into the cell. The net movement of positive ions into the postsynaptic cell causes a slight depolarization known as an excitatory postsynaptic potential (EPSP) (Figure 6.28). The EPSP is a depolarizing graded potential that decreases in magnitude as it spreads away from the synapse via local current. Its sole function is to bring the postsynaptic neuron’s membrane potential closer to threshold, facilitating action potential generation.
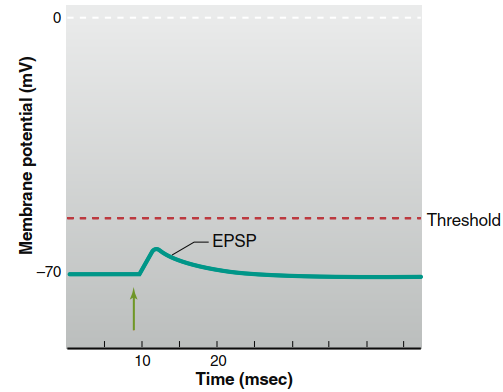
Figure 6.28. Excitatory postsynaptic potential (EPSP). Stimulation of the presynaptic neuron is marked by the green arrow. (Drawn larger than normal: typical EPSP 5 0.5 mV)
Inhibitory Chemical Synapses. At inhibitory chemical synapses, the postsynaptic potential change is typically a hyperpolarizing graded potential called an inhibitory postsynaptic potential (IPSP) (Figure 6.29). Alternatively, there may be no measurable IPSP but rather stabilization of the membrane potential at its existing value. In either scenario, activation of an inhibitory synapse reduces the likelihood that the postsynaptic cell will depolarize to threshold and generate an action potential.
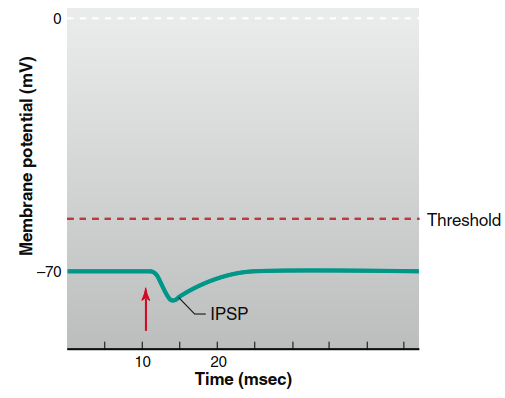
Figure 6.29. Inhibitory postsynaptic potential (IPSP). Stimulation of the presynaptic neuron is marked by the red arrow. (This hyperpolarization is drawn larger than a typical IPSP.)
At an inhibitory synapse, activated receptors on the postsynaptic membrane open Cl⁻ or K⁺ channels without affecting Na⁺ permeability. In cells that actively regulate intracellular Cl⁻ concentration via active transport out of the cell, the Cl⁻ equilibrium potential is more negative than the resting potential. Consequently, when Cl⁻ channels open, chloride enters the cell, producing hyperpolarization—an IPSP. In cells that do not actively transport Cl⁻, the equilibrium potential for chloride equals the resting membrane potential; increased Cl⁻ permeability does not change the membrane potential but enhances chloride’s influence, making it harder for excitatory inputs from other synapses to alter the potential when chloride channels are simultaneously open (Figure 6.30).
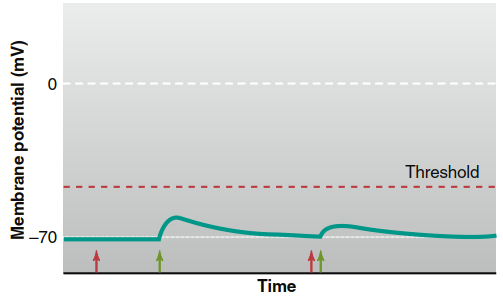
Figure 6.30. Synaptic inhibition of postsynaptic cells where ECl is equal to the resting membrane potential. Stimulation of a presynaptic neuron releasing a neurotransmitter that opens Cl- channels (red arrows) has no direct effect on the postsynaptic membrane potential. However, when an excitatory synapse is simultaneously activated (green arrows), Cl- movement into the cell diminishes the EPSP
Increased K⁺ permeability in the postsynaptic cell also produces an IPSP. If a cell membrane were permeable only to K⁺, the resting membrane potential would equal the K⁺ equilibrium potential (approximately -90 mV instead of the typical -70 mV). Thus, with increased K⁺ permeability, more potassium ions leave the cell, moving the membrane closer to the K⁺ equilibrium potential and causing hyperpolarization. This further reduces neuronal excitability, effectively inhibiting action potential generation.
Date added: 2026-07-14; views: 5;