Presynaptic and Postsynaptic Mechanisms of Synaptic Strength Modulation
Individual synaptic events, whether excitatory or inhibitory, are often presented as constant and reproducible, but enormous variability actually occurs in the postsynaptic potentials following a presynaptic input. The effectiveness or strength of a given synapse is influenced by both presynaptic and postsynaptic mechanisms. Understanding this variability is essential for explaining neuronal plasticity, learning, and drug actions.
Presynaptic Mechanisms. A presynaptic terminal does not release a constant amount of neurotransmitter every time it is activated. One major reason for this variation involves Ca²⁺ concentration. Calcium ions that enter the terminal during previous action potentials are pumped out of the cell or temporarily sequestered into intracellular organelles. If Ca²⁺ removal fails to keep up with entry—such as during high‑frequency stimulation—the Ca²⁺ concentration in the terminal increases, leading to greater neurotransmitter release upon subsequent stimulation. Consequently, more ion channels open in the postsynaptic membrane, amplifying the amplitude of the EPSP or IPSP.
The neurotransmitter output of some presynaptic terminals is also altered by activation of membrane receptors on the terminals themselves. Activation of these presynaptic receptors influences Ca²⁺ influx and thus the number of synaptic vesicles that release neurotransmitter into the cleft. These presynaptic receptors may be associated with a second synaptic ending known as an axo‑axonic synapse, where one neuron’s axon terminal ends on another’s axon terminal. For example, in Figure 6.33, the neurotransmitter released by neuron A binds to receptors on terminal B, changing the amount of neurotransmitter released from B in response to action potentials. Thus, neuron A has no direct effect on neuron C but exerts a presynaptic influence on the B‑to‑C synapse, either decreasing release (presynaptic inhibition) or increasing it (presynaptic facilitation).
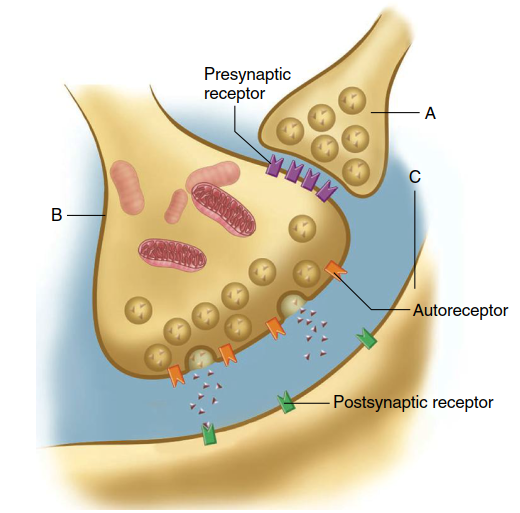
Figure 6.33. A presynaptic (axo–axonic) synapse between axon terminal A and axon terminal B. Cell C is postsynaptic to cell B
Axo‑axonic synapses, such as neuron A in Figure 6.33, can alter Ca²⁺ concentration in axon terminal B or even affect neurotransmitter synthesis there. The mechanisms vary across synapses: receptors on terminal B may be ionotropic, rapidly and directly changing the terminal’s membrane potential, or metabotropic, producing slower, longer‑lasting effects via second messengers. In either case, increased Ca²⁺ in terminal B raises the number of vesicles releasing neurotransmitter, while decreased Ca²⁺ reduces release. Axo‑axonic synapses are crucial because they selectively control specific inputs to the postsynaptic neuron C, particularly in sensory modulation, such as pain pathways (discussed in Chapter 7).
Some presynaptic receptors are not associated with axo‑axonic synapses. Instead, they are activated by neurotransmitters or other chemical messengers released by nearby neurons, glia, or even the axon terminal itself. In the last case, these receptors are called autoreceptors (see Figure 6.33) and provide negative feedback control, typically decreasing further neurotransmitter release. Autoreceptors thus allow a neuron to regulate its own output in response to changing activity levels.
Postsynaptic Mechanisms. Postsynaptic mechanisms also contribute to varying synaptic strength. As described in Chapter 5, multiple types and subtypes of receptors exist for each neurotransmitter. Different receptor types operate via distinct signal transduction mechanisms and can have different—sometimes opposite—effects on postsynaptic targets. A given transduction pathway may be regulated by several neurotransmitters, and different second‑messenger systems affecting the same ion channel can interact.
The number of receptors is not constant; it varies through up‑regulation and down‑regulation. Additionally, a receptor’s ability to respond to its neurotransmitter can change. In some systems, a receptor responds normally upon first exposure but eventually fails to respond despite continued presence of the neurotransmitter—a phenomenon called receptor desensitization. This desensitization partly explains why drug abusers develop tolerance to drugs that elevate certain brain neurotransmitters, requiring increasing doses to achieve the desired effect (see Chapter 8).
Further complexity arises when a cotransmitter (or several) is released alongside the primary neurotransmitter, acting on postsynaptic and possibly presynaptic receptors. Thus, even at a single synapse, the possible variations in transmission are immense, providing mechanisms for altering synaptic strength in response to changing conditions—a key aspect of plasticity introduced at the beginning of this chapter.
Modification of Synaptic Transmission by Drugs and Disease. The majority of therapeutic, illicit, and recreational drugs acting on the nervous system do so by altering synaptic mechanisms and thus synaptic strength. Drugs interfere with or stimulate normal processes involved in neurotransmitter synthesis, storage, release, and receptor activation. The synaptic mechanisms labeled in Figure 6.34 are vulnerable to drug effects.
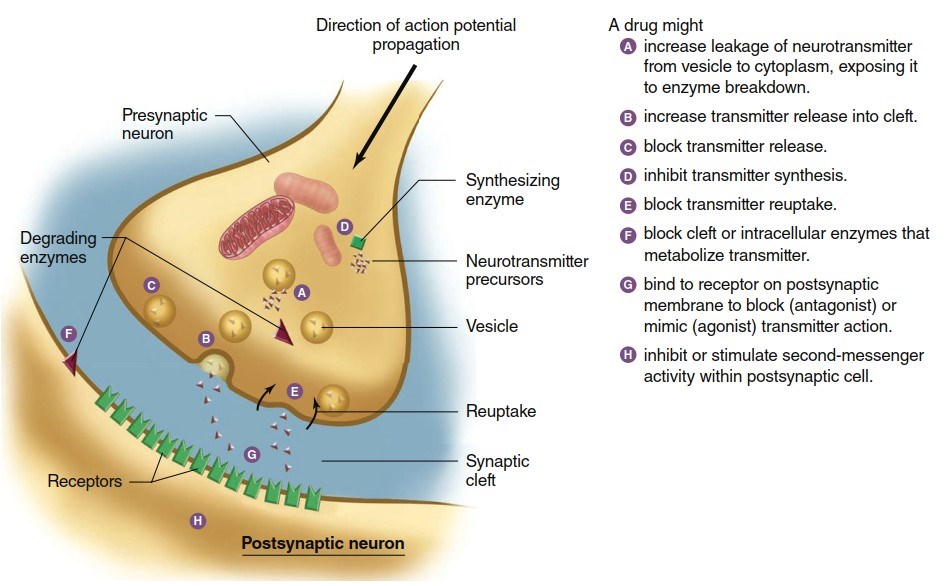
As introduced in Chapter 5, agonists are ligands that bind to a receptor and produce a response similar to normal activation, while antagonists bind but fail to activate, blocking the natural neurotransmitter. Specific agonists and antagonists can affect receptors on both presynaptic and postsynaptic membranes, providing targets for numerous medications and toxins.
Diseases also affect synaptic mechanisms. For instance, tetanus is caused by Clostridium tetani, which produces tetanus toxin—a protease that destroys SNARE proteins in the presynaptic terminal, preventing vesicle fusion and blocking neurotransmitter release. Tetanus toxin specifically affects inhibitory neurons in the CNS that normally suppress skeletal muscle activation. Consequently, tetanus toxin increases muscle contraction, causing rigid or spastic paralysis. In contrast, toxins from Clostridium botulinum (causing botulism) also block neurotransmitter release by destroying SNARE proteins but target excitatory synapses that activate skeletal muscles. Botulism therefore results in reduced muscle contraction, or flaccid paralysis. Low doses of one botulinum toxin (Botox) are used therapeutically for facial wrinkles, severe sweating, uncontrollable blinking, eye misalignment, and other conditions.

Table 6.5 summarizes the multiple factors that determine synaptic strength, integrating presynaptic release properties, receptor types, modulatory inputs, and disease mechanisms. Together, these presynaptic and postsynaptic variations enable the nervous system to adapt dynamically, yet they also render synapses vulnerable to pharmacological and pathological disruption.
Date added: 2026-07-14; views: 3;