Neuromodulators and Neurotransmitters: Mechanisms, Types, and Clinical Significance
Optimized Scientific Text. Researchers have traditionally emphasized the role of neurotransmitters in eliciting excitatory postsynaptic potentials (EPSPs) and inhibitory postsynaptic potentials (IPSPs). However, certain chemical messengers generate complex responses that cannot be classified simply as EPSPs or IPSPs. The term modulation describes these intricate responses, and the signaling molecules responsible are called neuromodulators. The distinctions between neuromodulators and neurotransmitters are not always clear-cut, as many neuromodulators are synthesized by the presynaptic cell and coreleased alongside classical neurotransmitters. Adding further complexity, numerous hormones, paracrine factors, and immune system messengers also function as neuromodulators.
Neuromodulators frequently modify the postsynaptic cell’s response to specific neurotransmitters, thereby amplifying or dampening the effectiveness of ongoing synaptic activity. Alternatively, they may alter the presynaptic cell’s synthesis, release, reuptake, or metabolic breakdown of a transmitter. In essence, neuromodulators change the overall efficacy of synaptic transmission rather than directly causing excitation or inhibition. This regulatory capacity allows neural circuits to adapt their output based on physiological demands and environmental cues.
In general, neurotransmitter receptors influence ion channels that directly affect postsynaptic cell excitation or inhibition through mechanisms operating within milliseconds. By contrast, neuromodulator receptors more commonly induce changes in neuronal metabolic processes, often via G proteins coupled to second-messenger systems. Such changes, which can unfold over minutes, hours, or even days, include alterations in enzyme activity and, through influences on DNA transcription, modifications in protein synthesis. Thus, neurotransmitters mediate rapid communication, while neuromodulators are associated with slower, sustained events such as learning, development, and motivational states.
The number of substances known to act as neurotransmitters or neuromodulators is large and continues to grow. Table 6.6 provides a framework for categorizing this list, though a comprehensive discussion exceeds the scope of this text. The following sections present only fundamental generalizations about key neurotransmitters, using the term neurotransmitter broadly while acknowledging that some messengers may be more appropriately described as neuromodulators. A note on terminology: neurons are often referred to using the suffix -ergic, where the missing prefix indicates the type of neurotransmitter released. For example, dopaminergic neurons release dopamine.

Acetylcholine (ACh). Acetylcholine (ACh) is a major neurotransmitter in the peripheral nervous system (PNS) at the neuromuscular junction, where a motor neuron contacts a skeletal muscle cell, and also in the brain. Neurons that release ACh are called cholinergic neurons. The cell bodies of the brain’s cholinergic neurons are concentrated in relatively few areas, but their axons are widely distributed throughout the central nervous system (CNS). This widespread projection pattern underlies acetylcholine’s diverse roles in attention, memory, and muscle control.
Acetylcholine is synthesized from choline—a common nutrient found in many foods—and acetyl coenzyme A within the cytoplasm of synaptic terminals, then stored in synaptic vesicles. After release and activation of postsynaptic receptors, the concentration of ACh at the postsynaptic membrane decreases due to the enzyme acetylcholinesterase, which stops receptor activation. This enzyme is located on both presynaptic and postsynaptic membranes and rapidly destroys ACh, releasing choline and acetate. The choline is then transported back into the presynaptic axon terminal for reuse in new ACh synthesis. Certain chemical weapons, such as the nerve gas Sarin, inhibit acetylcholinesterase, causing ACh buildup in the synaptic cleft, which leads to overstimulation of postsynaptic ACh receptors, initially causing uncontrolled muscle contractions and ultimately receptor desensitization and paralysis.
Nicotinic Acetylcholine Receptors. Although a receptor is considered specific for a given ligand such as ACh, most receptors recognize natural or synthetic compounds with some chemical similarity to that ligand. Some ACh receptors respond not only to acetylcholine but also to the compound nicotine and are therefore called nicotinic receptors. Nicotine is a plant alkaloid that constitutes 1% to 2% of tobacco products and is also contained in smoking cessation treatments such as nasal sprays, chewing gums, and transdermal patches. Nicotine’s hydrophobic structure allows rapid absorption through lung capillaries, mucous membranes, skin, and the blood-brain barrier.
The nicotinic acetylcholine receptor exemplifies a ligand-gated ion channel permeable to both sodium and potassium ions; however, because sodium has the larger electrochemical driving force, the net effect of channel opening is depolarization. Nicotinic receptors are present at the neuromuscular junction, and several nicotinic receptor antagonists are toxins that induce paralysis. In the brain, nicotinic receptors are important in cognitive functions and behavior; for instance, one cholinergic system using nicotinic receptors reinforces the ability to detect and respond to meaningful stimuli, playing a major role in attention, learning, and memory. The presence of nicotinic receptors on presynaptic terminals in reward pathways explains why tobacco products are among the most highly addictive substances.
Muscarinic Acetylcholine Receptors. The other general type of cholinergic receptor is stimulated not only by acetylcholine but also by muscarine, a poison found in certain mushrooms; these are consequently called muscarinic receptors. These receptors are metabotropic and couple with G proteins, which then alter the activity of various enzymes and ion channels. Muscarinic receptors are prevalent at some cholinergic synapses in the brain and at junctions where a major division of the PNS innervates peripheral glands, tissues, and organs such as salivary glands, smooth muscle cells, and the heart. Atropine is a naturally occurring antagonist of muscarinic receptors with many clinical uses, including eye drops that relax the smooth muscles of the iris, thereby dilating the pupils for ophthalmic examinations.
Alzheimer’s Disease. Many cholinergic neurons in the brain degenerate in individuals with Alzheimer’s disease, an age-related brain disease and the most common cause of declining intellectual function in late life. Alzheimer’s disease affects 10% to 15% of people over age 65 and 50% of people over age 85. Due to cholinergic neuron degeneration, this disease is associated with decreased ACh levels in certain brain areas and even loss of the postsynaptic neurons that would normally respond to ACh. These defects, along with abnormalities in other neurotransmitter systems, relate to the declining language abilities, cognitive dysfunction, confusion, and memory loss that characterize Alzheimer’s disease. Several genetic mechanisms contribute to increased risk; for example, a gene on chromosome 19 codes for a protein involved in cholesterol transport in the bloodstream.
Mutations of genes on chromosomes 1, 14, and 21 are associated with abnormally increased concentrations of beta-amyloid protein, which is linked to neuronal cell death in a severe form of the disease that can begin as early as age 30. Lifestyle factors such as diet, exercise, social engagement, and mental stimulation may also influence whether cholinergic neurons are lost and Alzheimer’s disease develops. Interestingly, synthetic chemicals that act like nerve gas but in a nontoxic manner are currently used to slow disease progression; these drugs do not restore lost cholinergic cells but increase acetylcholine concentration in synapses of remaining cells by inhibiting acetylcholinesterase.
Biogenic Amines. The biogenic amines are small, charged molecules synthesized from amino acids that contain an amino group (R—NH₂). The most common biogenic amines are dopamine, norepinephrine, serotonin, and histamine. Epinephrine, another biogenic amine, is not a common neurotransmitter in the CNS but is the major hormone secreted by the adrenal medulla. Norepinephrine is an important neurotransmitter in both the central and peripheral components of the nervous system, regulating arousal, blood pressure, and mood.
Catecholamines. Dopamine (DA), norepinephrine (NE), and epinephrine all contain a catechol ring (a six-carbon ring with two adjacent hydroxyl groups) and an amine group, which is why they are called catecholamines. Catecholamines are formed from the amino acid tyrosine and share the same two initial steps in their synthetic pathway (Figure 6.35). Synthesis begins with tyrosine uptake by axon terminals and its conversion to L-dihydroxy-phenylalanine (L-dopa) by the rate-limiting enzyme tyrosine hydroxylase. Depending on the enzymes expressed in a given neuron, any one of the three catecholamines may ultimately be released. Autoreceptors on presynaptic terminals strongly modulate catecholamine synthesis and release, providing feedback regulation.
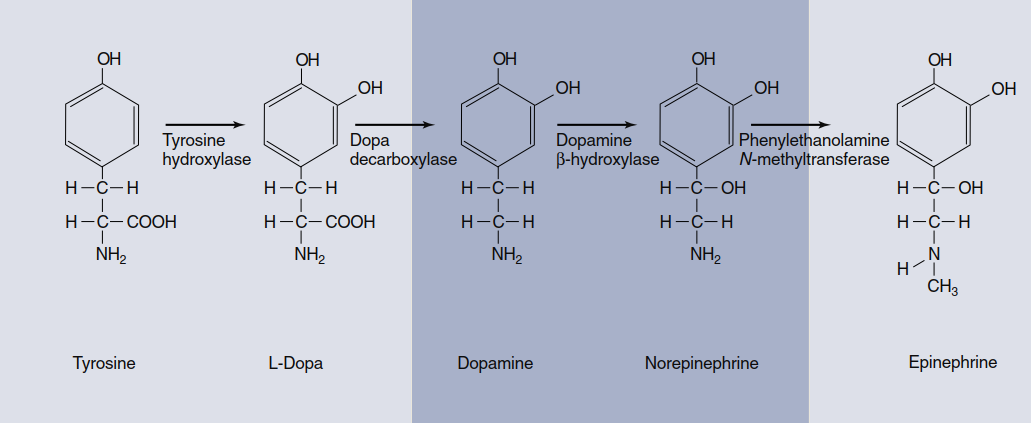
Figure 6.35. Catecholamine biosynthetic pathway. Tyrosine hydroxylase is the rate-limiting enzyme, but which neurotransmitter is ultimately released from a neuron depends on which of the other three enzymes are present in that cell. The dark-colored box indicates the more common CNS catecholamine neurotransmitters. Epinephrine is primarily a hormone released by the adrenal glands.
After activation of postsynaptic receptors, catecholamine concentration in the synaptic cleft declines mainly because a membrane transporter protein actively transports the catecholamine back into the axon terminal. Catecholamine neurotransmitters are also broken down in both the extracellular fluid and the axon terminal by enzymes such as monoamine oxidase (MAO). Drugs known as monoamine oxidase (MAO) inhibitors increase norepinephrine and dopamine levels in synapses by slowing their metabolic degradation and are used in treating mood disorders such as certain types of depression. Within the CNS, catecholamine-releasing neuron cell bodies lie in the brainstem and hypothalamus; although these neurons are relatively few, their axons branch extensively and project to virtually all parts of the brain and spinal cord. These neurotransmitters have essential functions in states of consciousness, mood, motivation, directed attention, movement, blood pressure regulation, and hormone release.
Epinephrine and norepinephrine are also synthesized in the adrenal glands. For historical reasons related to nineteenth-century physiologists referring to adrenal secretions as “adrenaline,” the adjective adrenergic describes neurons that release norepinephrine or epinephrine and also the receptors to which these neurotransmitters bind. There are two major receptor classes for norepinephrine and epinephrine: alpha-adrenergic receptors (alpha-adrenoceptors) and beta-adrenergic receptors (beta-adrenoceptors). All catecholamine receptors are metabotropic and use second messengers to transfer signals from the cell surface to the cytoplasm. Alpha-adrenoceptors exist in two subclasses, α₁ and α₂; they act presynaptically to inhibit norepinephrine release (α₂) or postsynaptically to stimulate or inhibit different types of K⁺ channels (α₁). Beta-adrenoceptors act via stimulatory G proteins to increase cAMP in the postsynaptic cell. There are three beta-receptor subclasses—β₁, β₂, and β₃—which function differently in various tissues. These subclasses are distinguished by the drugs that influence them and their second-messenger systems.
Serotonin. Serotonin (5-hydroxytryptamine, or 5-HT) is produced from tryptophan, an essential amino acid. Its effects generally have a slow onset, indicating that it works primarily as a neuromodulator. Serotonergic neurons innervate virtually every structure in the brain and spinal cord and operate via at least 16 different receptor subtypes, reflecting serotonin’s wide-ranging modulatory influence. In general, serotonin has an excitatory effect on pathways involved in muscle control and an inhibitory effect on pathways that mediate sensory information. The activity of serotonergic neurons is lowest or absent during sleep and highest during states of alert wakefulness. In addition to contributions to motor activity and sleep, serotonergic pathways regulate food intake, reproductive behavior, and emotional states such as mood and anxiety.
Selective serotonin reuptake inhibitors (SSRIs) such as paroxetine (Paxil) aid in treating depression by inactivating the presynaptic membrane 5-HT transporter that mediates serotonin reuptake into the presynaptic cell, thereby increasing synaptic neurotransmitter concentration. Interestingly, such drugs are often associated with decreased appetite but paradoxically cause weight gain due to disruption of enzymatic pathways that regulate fuel metabolism. This example illustrates how reuptake inhibitors for a neurotransmitter with widespread actions can produce unwanted side effects. Serotonin is found in both neural and nonneural cells, with the majority located outside the CNS; approximately 90% of the body’s total serotonin is in the digestive system, 8% in blood platelets and immune cells, and only 1% to 2% in the brain. The drug lysergic acid diethylamide (LSD) stimulates the 5-HT₂A subtype of serotonin receptor in the brain, and alteration of this receptor complex produces the intense visual hallucinations characteristic of LSD ingestion.
Amino Acid Neurotransmitters. In addition to neurotransmitters synthesized from amino acids, several amino acids themselves function directly as neurotransmitters. Although amino acid neurotransmitters chemically fit the category of biogenic amines, they are traditionally placed in their own category. Amino acid neurotransmitters are by far the most prevalent neurotransmitters in the CNS and affect virtually all neurons there.
Glutamate. Among the excitatory amino acids, the most common by far is glutamate, estimated to be the primary neurotransmitter at 50% of excitatory synapses in the CNS. Pharmacological manipulation of glutamate receptors has permitted identification of specific receptor subtypes by their ability to bind natural and synthetic ligands. Although metabotropic glutamate receptors exist, the vast majority are ionotropic, with two important subtypes found in postsynaptic membranes: AMPA receptors (identified by binding to α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid) and NMDA receptors (which bind N-methyl-D-aspartate).
Cooperative activity of AMPA and NMDA receptors is implicated in long-term potentiation (LTP) , a mechanism that couples frequent synaptic activity with lasting changes in signaling strength across that synapse, thought to be a major cellular process underlying learning and memory. Figure 6.36 outlines the mechanism stepwise. When a presynaptic neuron fires action potentials (step 1), glutamate is released from presynaptic terminals (step 2) and binds to both AMPA and NMDA receptors on postsynaptic membranes (step 3). AMPA receptors function like typical excitatory postsynaptic receptors: when glutamate binds, the channel becomes permeable to both Na⁺ and K⁺, but the larger Na⁺ entry creates a depolarizing EPSP (step 4). By contrast, NMDA receptor channels also mediate substantial Ca²⁺ flux, but opening them requires more than glutamate binding. A magnesium ion blocks NMDA channels when the membrane voltage is near the negative resting potential; to drive it out, the membrane must be significantly depolarized by current through AMPA channels (step 5). This explains why high-frequency presynaptic action potentials are required to complete LTP. At low frequencies, insufficient temporal summation of AMPA-receptor EPSPs prevents the 20–30 mV depolarization needed to displace magnesium, so NMDA receptors do not open. When depolarization is sufficient, NMDA receptors open, allowing Ca²⁺ to enter the postsynaptic cell (step 6).
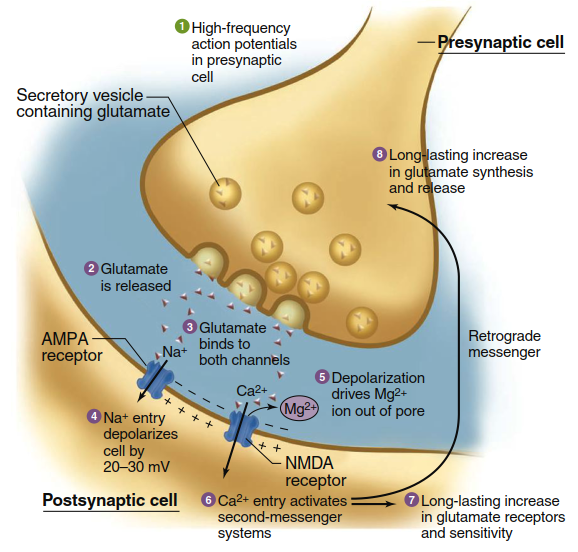
Figure 6.36. Long-term potentiation at glutamatergic synapses. Episodes of intense firing across a synapse result in structural and chemical changes that amplify the strength of synaptic signaling during subsequent activation. See text for description of each step; details of the mechanism linking steps 1 and 2 were described in Figure 6.27. Note that both AMPA and NMDA receptors are nonspecific cation channels that also allow K1 flux, but the net Na1 and Ca21 fluxes indicated are most relevant to the LTP mechanism, as described in the text.
Calcium ions then activate a second-messenger cascade including persistent activation of multiple protein kinases, stimulation of gene expression and protein synthesis, and ultimately a long-lasting increase in postsynaptic sensitivity to glutamate (step 7). This system can also activate long-term enhancement of presynaptic glutamate release via unidentified retrograde signals (step 8). Each subsequent action potential along this presynaptic cell causes greater postsynaptic depolarization, so repeatedly activating a particular synaptic pattern causes chemical and structural changes that facilitate future activity along those same pathways.
NMDA receptors have also been implicated in excitotoxicity, a phenomenon where injury or death of brain cells (e.g., from blocked or ruptured blood vessels) rapidly spreads to adjacent regions. When glutamate-containing cells die and their membranes rupture, the glutamate flood excessively stimulates AMPA and NMDA receptors on nearby neurons. Excessive stimulation causes toxic concentrations of intracellular Ca²⁺, killing those neurons and causing them to rupture, progressively spreading damage. Recent experiments and clinical trials suggest that administering NMDA receptor antagonists may help minimize the spread of cell death following brain injuries.
GABA. GABA (gamma-aminobutyric acid) is the major inhibitory neurotransmitter in the brain. Although not one of the 20 amino acids used to build proteins, it is classified with amino acid neurotransmitters because it is a modified form of glutamate. With few exceptions, GABA neurons in the brain are small interneurons that dampen activity within neural circuits. Postsynaptically, GABA may bind to ionotropic or metabotropic receptors. The ionotropic receptor increases Cl⁻ flux into the cell, resulting in hyperpolarization (an IPSP) of the postsynaptic membrane. In addition to the GABA binding site, this receptor has several additional binding sites for other compounds, including steroids, barbiturates, and benzodiazepines. Benzodiazepine drugs such as alprazolam (Xanax) and diazepam (Valium) reduce anxiety, guard against seizures, and induce sleep by increasing Cl⁻ flux through the GABA receptor.
Synapses using GABA are also among the many targets of ethanol (ethyl alcohol) in alcoholic beverages. Ethanol stimulates GABA synapses and simultaneously inhibits excitatory glutamate synapses, with the overall effect being global depression of brain electrical activity. Thus, as blood alcohol content increases, there is progressive reduction in overall cognitive ability, along with sensory perception inhibition (particularly hearing and balance), loss of motor coordination, impaired judgment, memory loss, and unconsciousness. Very high ethanol doses can be fatal due to suppression of brainstem centers regulating circulatory and respiratory systems. Dopaminergic and endogenous opioid signaling pathways are also affected by ethanol, resulting in short-term mood elevation or euphoria, and involvement of these pathways underlies long-term alcohol dependence in some individuals.
Glycine. Glycine is the major neurotransmitter released from inhibitory interneurons in the spinal cord and brainstem. It binds to ionotropic receptors on postsynaptic cells that allow Cl⁻ to enter, preventing these cells from approaching the threshold for firing action potentials. Normal function of glycinergic neurons is essential for maintaining a balance of excitatory and inhibitory activity in spinal cord integrating centers that regulate skeletal muscle contraction. This balance becomes apparent in poisoning with the neurotoxin strychnine, an antagonist of glycine receptors sometimes used to kill rodents. Affected individuals experience hyperexcitability throughout the nervous system, leading to convulsions, spastic contraction of skeletal muscles, and ultimately death due to impairment of respiratory muscles.
Neuropeptides. Neuropeptides are composed of two or more amino acids linked by peptide bonds. Approximately 100 neuropeptides have been identified, though their physiological functions are not all known. Evolution has favored the same chemical messengers for use in widely differing circumstances, and many neuropeptides were previously identified in nonneural tissue where they function as hormones or paracrine substances; they generally retain the names given upon their initial discovery in nonneural tissue.
Neuropeptides are formed differently from other neurotransmitters, which are synthesized in axon terminals by few enzyme-mediated steps. Neuropeptides, in contrast, are derived from large precursor proteins that themselves have little inherent biological activity. Synthesis of these precursors, directed by mRNA, occurs on ribosomes located only in the cell body and large dendrites of the neuron, often a considerable distance from axon terminals or varicosities where peptides are released. In the cell body, the precursor protein is packaged into vesicles, which are then moved by axonal transport to terminals or varicosities (review Figure 6.3), where the protein is cleaved by specific peptidases. Many precursor proteins contain multiple peptides, which may be different or copies of one peptide. Neurons that release one or more peptide neurotransmitters are collectively called peptidergic. In many cases, neuropeptides are cosecreted with another neurotransmitter type and act as neuromodulators.
The amount of neuropeptide released from vesicles at synapses is significantly less than that of nonpeptidergic neurotransmitters such as catecholamines. Additionally, neuropeptides can diffuse away from the synapse and affect distant neurons, in which case they are referred to as neuromodulators. These actions last longer (on the order of several hundred milliseconds) than when neuropeptides or other molecules act as neurotransmitters. After release, neuropeptides can interact with either ionotropic or metabotropic receptors and are eventually broken down by peptidases located in neuronal membranes.
Endogenous opioids—a group of neuropeptides including beta-endorphin, the dynorphins, and the enkephalins—have attracted much interest because their receptors are the sites of action of opiate drugs such as morphine and codeine. Opiate drugs are powerful analgesics (pain relievers without loss of consciousness), and endogenous opioids undoubtedly function in pain regulation. Evidence also indicates that opioids regulate eating and drinking behavior, circulatory system function, mood, and emotion.
Gases as Neurotransmitters. Certain very short-lived gases also serve as neurotransmitters. Nitric oxide is the best understood, but recent research indicates that carbon monoxide and hydrogen sulfide are also emitted by neurons as signals. Gases are not released by exocytosis of presynaptic vesicles, nor do they bind to postsynaptic plasma membrane receptors. They are produced by enzymes in axon terminals (in response to Ca²⁺ entry) and simply diffuse from their sites of origin in one cell into the intracellular fluid of other neurons or effector cells, where they bind to and activate proteins. For example, nitric oxide released from neurons activates guanylyl cyclase in recipient cells, increasing the concentration of the second messenger cyclic GMP, which can then alter ion channel activity in the postsynaptic cell.
Nitric oxide functions in a wide array of neurally mediated events, including learning, development, drug tolerance, penile and clitoral erection, and sensory and motor modulation. Paradoxically, it is also implicated in neural damage resulting from stoppage of blood flow to the brain or from head injury. Nitric oxide is produced not only in the central and peripheral nervous systems but also by various nonneural cells; for example, it has important paracrine functions in the circulatory and immune systems, among others.
Purines as Neuromodulators. Other nontraditional neurotransmitters include the purines, ATP and adenosine, which act principally as neuromodulators. ATP is present in all presynaptic vesicles and is coreleased with one or more other neurotransmitters in response to Ca²⁺ influx into the terminal. Adenosine is derived from ATP via enzyme activity in the extracellular compartment. Both presynaptic and postsynaptic receptors have been described for adenosine, and the functions of these substances in the nervous system and other tissues remain active areas of research.
Date added: 2026-07-14; views: 3;