Effectiveness of Dispersal Mechanisms
Dispersal can be limited by physical barriers, such as large differences in altitude of mountainous regions, oceans and large arid regions. The absence of certain environmental conditions or requirements (i.e. temperature, water, light, soil nutrients, symbiotic patterns) may significantly limit successful establishment following dispersal. Keep in mind that these barriers or requirements can be manipulated in plants favoured by the human vector, where people can improve growing conditions by removing competition, improving soil conditions through fertilisation or aeration.
A clear understanding of how far propagules can disperse is especially important in the field of ecosystem restoration, which could provide key information about species’ ability to persist in fragmented habitats. As this fragmentation is quite common in human-altered landscapes, better knowledge about successful seed dispersal is indispensable. This is especially true for ballistic and other autochoric species, including ant- and bat-dispersed vectors, where dispersal distances travel is rarely beyond a few metres. In contrast, many vertebrates, wind and water are able to disperse propagules for distances exceeding 10 km. In some of the best-case dispersal distance scenarios, orchid seeds and spores from ferns have been known to travel distances of several hundred kilometres.
When flora becomes established on young islands or within post-disturbance landscapes, it will occur primarily by anemochoric species, unless a human vector facilitates its introduction. Once established, primary dispersal mechanisms tend to shift from primarily wind to water or birds, as illustrated in a fallow field (Fig. 18.2). During the first few years, wind-dispersed species become established predominantly via longdistance transport; however, as time passes, allochoric species become more dominant after the first two decades and, finally, typically within a century, replace wind-dispersed species. Other quantitative aspects of seed dispersal to consider include (1) the number of seeds produced and dispersed, (2) the number of visits of dispersers (e.g. birds), and (3) the continuity of favourable weather conditions (e.g. duration of air flow). The efficiency of seed dispersal also has qualitative aspects. The quality of a seed may determine whether or not a bird will decide to disperse it, how the bird manipulates it and, finally, how it is transported, all of which will affect the probability of seed germination.
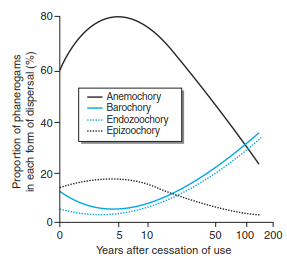
Fig. 18.2. Dispersal mechanisms can change during succession. Relative frequency of four dispersal mechanisms in phanerogamic flora of fallow fields close to Montpellier (France) at different times after their abandonment (after Lepart and Escarre 1983)
The dispersal of propagules is usually deemed successful if they reach a site that is favourable for their germination and establishment and ensures growth till flowering and seed production (safe site). Following germination and establishment it is extremely important to the long-term survival of the species that it exploit its new site, which is at some distance from the parent plant, contributing to the expansion of the species range or area. An expanding range or shift provides a few benefits, especially if it occurs at fast enough rates. A fast shift can allow a plant species to escape certain pathogens and enhance the population’s chances of survival.
To a large degree this depends on the available mechanisms for dispersal and their effectiveness regarding a targeted (suitable site for growth) and far-reaching (gain space) transport of propagules. However, such substrate “targets” are typically not achieved through a single form of dispersal. To ensure successful colonisation, a series of different dispersal vectors and their associated interactions can be used to improve seed dispersal (heterospory). Multiple dispersal mechanisms can supplement each other to ensure successful dispersal. Under unfavourable external conditions (i.e. a short vegetation period in high mountains or long dry periods), reproductive dispersal is combined with vegetative propagation. For example, the alpine species Polygonum viviparum produces both seeds and bulbils on the same stalk, with the bulbil fraction increasing with altitude.
These particular advantages associated with specific regions typically occur when the species adapts to the local conditions over time (local adaptation). For instance, wind-dispersed propa- gules in arid regions are often transported over long distances because only minimal obstacles occur over land. Another example of plant- dispersal local adaption is the production of flashy or tasty fruit-bearing plants, which increase the likelihood of attracting animal dispersal vectors. Essentially, different strategies have evolved in which successful arrival at safe sites and spatial expansion provide a niche balance. Studies on seed dispersal in forests have shown that many trees in the humid tropics with rainy and dry seasons have created morphologically adapted seeds that facilitate transportation by animals. In temperate forests this also applies to shrubs, but here seeds from trees are predominantly transported by wind. This change in the form of dispersal is explained by regular winds occurring in temperate latitudes. Also in dry areas, where almost no obstacles impede air currents, wind dispersal dominates. All this is to say that there is no concrete evidence for certain dispersal vectors being exclusively used or associated with specific biomes (Tables 18.1 and 18.2).
Several hypotheses and models have been developed to help explain the link between certain plant species and their vectors. The low investment model is associated with plants’ investing little per propagule produced, resulting in a large number of small seeds. This model favours dispersal vectors associated with longdistance dispersal (wind-dispersed, attached to animals). Because little is invested in each seed, the recruitment rate is quiet low; however, this can be offset by volume. This strategy is typically used by early successional species. The second model, the high investment model, is essentially the opposite, where plants produce very few, large seeds. The large seed size provides ample amounts of energy and nutrients to ensure good rates of germination, assuming it attracts the appropriate dispersing agent (bird or bat) and falls upon a favourable substrate.
The seeds in this strategy are only distributed over short distances, but they usually find relatively safe conditions for germination. In animal-dispersed species, both strategies are used: seeds externally attached to animals (with low investment) and those transported in the gut (with high investment). The directed dispersal hypothesis first proposed by Howe and Smallwood (1982), regarding targeted, relatively safe dispersal, also aligns with the high investment model. This contrasts with the colonisation hypothesis, where opportunists exploit opportunities for rapid dispersal over large areas. The premise for the escape hypothesis is that the chances for establishment and germination depend on the low density of conspecifics, so the mother plant cannot be nearby (e.g. Janzen-Connell effect) (Sect. 19.3).
Only a few experimental results have confirmed these particular hypotheses. It is certain, however, that many plants do not use only one vector but possess morphological and other adaptations to exploit several vectors. Such polychory improves successful dispersal considerably. The most important components for efficient seed dispersal are summarised in a hierarchical classification (Table 18.3).
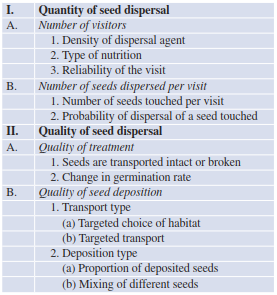
Table 18.3. Factors affecting efficiency of dispersal (after Schupp 1993)
Date added: 2026-04-26; views: 151;