Vegetation Geography. Characterisation and Interpretation of Areas
One consequence of short-range dispersal is that the site where propagules fall to the ground is already occupied by conspecific plants, often offspring of the same mother plant. Hence, growing seedlings will typically face intraspecific competition because a favourable abiotic environment is likely. In contrast, for propagules transported by long-distance dispersal, there is typically a higher chance of facing unfavourable abiotic and biotic factors. Although the likelihood of establishment at less favourable sites is lower, a successful establishment can still lead to spatial expansion. The result is an increased species range or area, which is a geographically defined region where individuals of a species can be found.
The science of spatial distribution of plants (chorology) aims (1) to recognise the various types and characteristic patterns of distribution, which provide the ability to map and to describe species organisation, and (2) to explain the development of patterns of distribution. A comparative assessment of geographical areas leads to the characterisation of floristic elements and areal types or geo-elements.
Characterisation and Interpretation of Areas.There are various ways to map areas. Frequently, when considering species distributions, only borders are drawn without providing any additional information concerning the distribution and abundance of species within the area. This can be avoided by creating dot maps, which can provide species locations. The most popular way to represent species distributions is through the use of species range maps. However, these maps often show only a portion of the entire range and rarely contain any information about species abundance. Such maps also show the climatic differentiation of areas often better than those based on meteorological data. Regardless of their shortcomings, these maps are indispensable for applied tasks, for example, nature protection.
The species range or area is essentially the spatial distribution in which a biological taxon is found, that is, where the environment is typically favourable for that species. The range limits are typically controlled by a number of historical and ecological factors, for example, climate, interspecific (other species) and intraspecific (same species) competition, site quality, food resources, water and landscape. Isolating the contribution of each of these factors individually is extremely difficult, especially since they can be codependent on each other. One attempt at determining range limits has been done through the use of isotherms (average temperatures); however, plants rarely react to average conditions. For example, the eastern species range of beech (Fagus sylvatica) is constrained by the lack of precipitation; however, another constraint is the severe cold winter (> -30 °C), which is known to cause extensive damage to bud development.
At the species’ southern range limit, the opposite occurs, where a combination of a lack of summer precipitation and high temperatures causes significant drought events. Finally, beech’s northern limit has been determined by a combination of prolonged winters and late-season frost events. However, it has been speculated that beech reached its northern limit in the north-west of the British Isles after the last glaciation. These observations have relied on the current distribution patterns of adult trees while ignoring environmental constraints on seed germination and seedling establishment, key processes in determining any long-term presence of a species. This will be of special importance in studies about climate-change-driven dynamics as species attempt to maintain their climatic niches by expanding their ranges in altitude and latitude. As decisive for alpine plant communities and their upper distribution boundary is summer frost resistance (Taschler and Neuner 2004).
Identifying the constraining factors influencing range limit is particularly important for olive trees in the Mediterranean region (Fig. 18.3). In the mountainous portion of these regions, prolonged frost, coupled with a shorter growing season, minimises biomass accumulation in the stem. In southern Europe, early summer is often too moist, inhibiting pollination and making fruit setting difficult. In North Africa, towards the south, increasing drought is limiting. At these limits, Walter’s law of the relative constancy of habitats can be applied to understand the occurrence of plants. In the southern species range of Quercus pubescens, for example, its occurrence indicates moist outposts of Mediterranean habitats, while at the northern range, in the upper Rhine Valley, it is found only on the warmest and driest, south-facing slopes. If the climate within the area of a plant species is changing in a particular direction, this species moves into a habitat that compensates for this climate change, so that the environmental conditions remain rather constant, assuming that the demands of a specific plant species remain constant (Walter 1986).
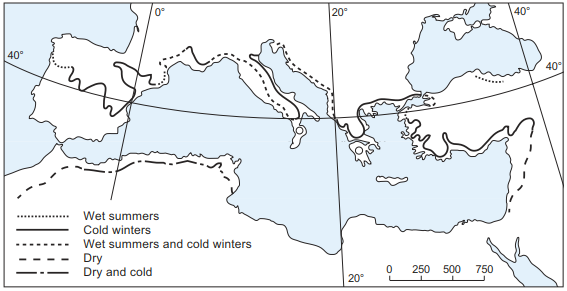
Fig. 18.3. Limit of the distribution of olive trees (Olea europaea) in the Mediterranean area and the most important climatic factors determining it (after Müller-Hohenstein 1981)
It should be mentioned that species range limits are dynamic, under constant flux, and can change quite readily owing to climatic factors influencing the biological system. The postglacial retreat of ice caused by climate warming led to significant species range shifts, which made it possible to make inferences on the rates of possible migration. For some important Central European tree species a range expansion of approximately 100 m/year was calculated, a considerable rate when one considers that trees are sessile organisms that typically require decades to reach reproductive maturity. Furthermore, quiet often many tree species will produce ample seed crops only once every several years (masting).
Considering the changes associated with climate change, species invasion, natural disturbances and new land-use practices, inventorying species ranges will continue. Complicating matters, the presence of synanthropic species or species that benefit from the presence of humans makes mapping species’ ranges and range limits extremely difficult. Ecologists have thus introduced the concept of a potential or fundamental range and a realised range. A potential range is an area where the range of environmental conditions is suitable for existence without the influence of competition or predation from interspecific species, whereas a realised range is the part of the potential range that is actually occupied by the species. A realised range is typically limited by various site conditions. Figure 18.4 shows schematically how differently the spatio-temporal formation of areas may occur. Areas may grow or shrink; they can also divide (disjunction), and new taxa may evolve by allopatric differentiation (metapopulations of the same species live in clearly separated geographical areas).
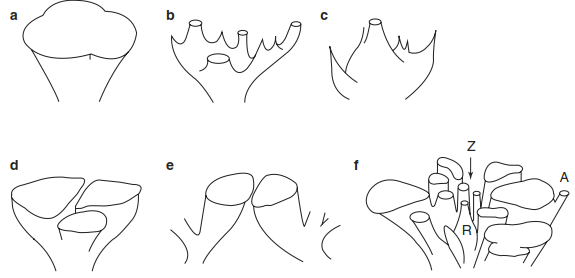
Fig. 18.4. Possible means by which plants exploit space over a period of time. Time axis vertical, space axis horizontal, cut surfaces show the current situation. a Area expansion; b death of populations and shrinkage to disjunct areas; c as in b with development of relict palaeoendemic areas; d allopatric differentiation of three vicarious related groups; e pseudovicarious species, living in ecologically or geographically similar conditions; f distribution of closely related species with a centre of diversity (Z), relict endemics (R) and neoendemic species (A). There are no fixed relationships between the age of the species, diversity of the group or size of the area (from Strasburger and Sitte 1998)
Species ranges are also characterised by their (1) size, (2) form and (3) geographical location. The differentiation between cosmopolitan and endemic species is based on the size of distribution areas. Cosmopolitan species typically occur over large areas, commonly extending over continents and different climatic zones. These species have extremely effective dispersal mechanisms, are highly competitive and can be categorised as phylogenetically old. Some common examples of cosmopolitan species include bracken (Pteridium aquilinum) and annual meadow grass (Poa annua). Cosmopolitans are differentiated from species that are generalists in terms of their site requirements (e.g. Pinus sylvestris).
Endemic species are species unique to one specific location or distribution, for example, in isolated mountain regions or islands. Two types of endemic species exist: palaeo-endemic and neo-endemic. A palaeo-endemic (sometimes referred to as relict-endemic) species is a species that historically was known to be widely distributed but, owing to either a disturbance (natural or anthropogenic) or competition, is now restricted to a much smaller native area. Typically, these species are phylogenetically old species. The giant redwoods (Sequoia gigantea) of California and the Eastern Asian Ginkgo biloba are both classic examples of these types of species. In contrast, neo-endemics, the phylogenetically younger counterpart, are species that have not had the opportunity temporally to benefit from inhabiting a larger range. Another reason for their lack of distribution could be attributed to physical barriers in the landscape, which has limited such movement. Island biogeographers have collected data from island groups that have shown that the distance of an island to the mainland, coupled with the duration of isolation, reflects the proportion of endemics in flora composition (Table 18.4).
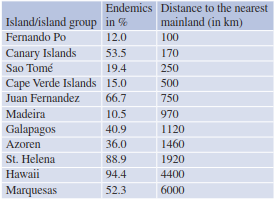
Table 18.4. Proportion of endemics in the flora of different islands and island groups (after Frey and Lösch 1998)
Species areas are also differentiated by their form, particularly whether they are closed areas, that is, the species is established in a single, clearly delimited space, or whether the area consists of several partial areas and is thus disjunct. In closed areas, gaps between individual growing sites are so small that they may be bridged easily and quickly by the transport of propagules or pollen. In disjunct areas, this is no longer possible. It may be assumed that polyphyletic origin of the same species does not occur and therefore other explanations for the genesis of disjunct areas must be found. One explanation might be an extremely rare distribution event, for example, by migrating birds or through some anthropogenic influence. In many cases, however, it is known that present disjunct areas were once closed and were separated by tectonic events (e.g. continental drift), the formation of mountains or climatic changes (e.g. change in cold and warm periods). A good example of the latter are arcto- alpine species, which were widely distributed in Europe during the Pleistocene, growing in the lowland tundra habitats between the Scandinavian and alpine glaciers. Today, under a warmer period, they are restricted to high alpine and arctic habitats, as well as to isolated patches in lower mountain ranges in Central Europe (e.g. Dryas octopetala).
With the separation of once closed areas, so- called vicarious areas may develop. In these partial areas, the populations of once uniform species developed further in different ways. This would be called geographical vicariance with examples of the genera Kleinia and Aeonium (Fig. 18.5). The centres of diversity of these species in southern and eastern tropical Africa as well as in Macaronesia (including the Cape Verde islands) are separated by the North African dry regions. If subspecies or other species of a genus are established in the same space at different sites, this is called ecological vicari- ance; examples are the two alpine rhododendrons Rhododendron hirsutum and R. ferrugineum, respectively, on soils of limestone and silicate rocks.
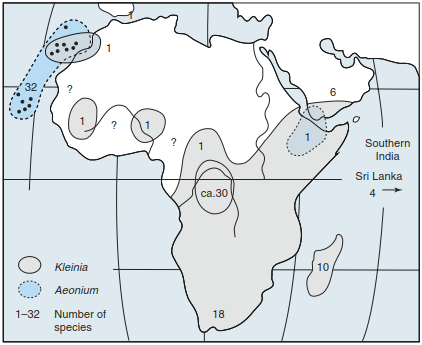
Fig. 18.5. Vicariance of the two genera Kleinia and Aeonium (Deil and Müller-Hohenstein 1984, after Richter 1997)
In general terms, differences in geological site conditions are related to differences in flora. The question of what factors cause these changes in a flora can only be answered at the level of the ecosystem. For example, on siliceous and limestone bedrock, the following ecological conditions of the site are altered:
- Limestone sites are usually drier than siliceous sites, because the water from precipitation seeps rapidly into deep soil layers.
- Limestone sites are warmer than siliceous sites because of the lower soil water content.
- Siliceous: deficient in Mo and alkaline cations but with an excess of Al.
- On limestone sites litter decomposition leads to mulch (mull) as the dominating form of humus, while siliceous sites are characterised by raw humus or moder.
- N mineralisation on limestone sites leads to nitrate as the dominating N form, whereas on siliceous sites it is ammonium.
The multitude of factors affecting a site shows that there is not a single factor that explains differences in vegetation. Most plants growing on siliceous substrates do not grow on limestone because of the soil chemistry (Fig. 18.6). Conversely, competition in the root layer stops the immigration of limestone species into siliceous sites, although they could grow there without competition. While 34 species of the siliceous matgrass meadows (Nardetum) are not able to colonise calcareous bluegrass meadows (Seslerietum) because of abiotic factors such as Fe deficiency, a large supply of phosphate or Al toxicity, it is mainly competition by roots of the matgrass species that stops invasion by bluegrass species. These abiotic factors, on the other hand, only affect nine species (Gigon 1987).
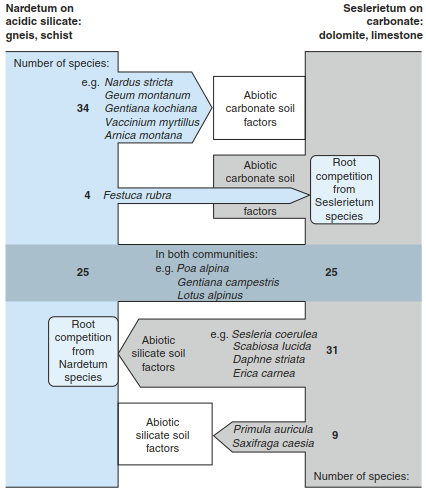
Fig. 18.6. Relationship between calcareous and siliceous vegetation of two alpine plant ecosystems. Matgrass communities (Nardus stricta) occur on siliceous sites, and bluegrass meadows (Sesleria coerulea) occupy calcareous sites. The number of species unable to invade the other’s root space, either because of root competition or other abiotic factors, is shown (after Gigon 1987)
If phylogenetically related species are distributed closely together, it may be concluded that they have developed within the same region and that the area is a centre of diversity (genetic centre, central zone of related groups), or at least a maintenance centre of this genus. Plant breeders aim to find such centres for economically important plants because they hope to find important gene reserves.
Date added: 2026-04-26; views: 164;