Species-Area Relationships. Equilibrium Theory of Island Biogeography
The spatial distribution of co-occurring plants depends on the dispersal mechanisms of species, rates of reproduction, competitiveness, growth and other factors. In nature, clumped distributions are particularly frequent, rather “island-like”, where patches are covered more densely, so that there are differences in abundance. Ecologists seeking the origin of such patterns need to know how many species (and individuals) are able to live in a certain area, known as the so-called species-area relationship (SAR).
Species-area relationships are an important background for understanding biodiversity. No single factor determines these relationships, and our present knowledge about how many factors there are and how they interact (e.g. random placement, minimum area effects and evolutionary independence) remains limited. Incomplete surveys in heterogeneous habitats contribute to the present difficulties in formulating a clear definition of these relationships, where several types currently exist. Further, attempts at clarifying species-area relations have sometimes been based on a rather small empirical basis for mathematical modelling. While mathematical models provide some insight into these interactions, they usually are based on initially simple equations, reflecting reality to only some degree, sometimes leading to incorrect conclusions. In contrast, holistic attempts have also been limited in their success, which have often led to superficial descriptive conclusions.
In this complex context it is most important to consider the spatial scales, which may differ with respect to the importance of the interactions that occur. The variability of scale is vast, from tiny soil crusts in an extreme desert to large species-rich tropical forests. Since long-term global vegetation surveys are well established— especially in the context of the relationship between climate and vegetation—large-scale maps and graphs are widely used and generally well accepted (Koppen 1900; Holdridge 1966). In these examples, vegetation is based on a classification of vegetation formations, such as evergreen rainforest, dry savanna and desert shrubland, which are characterised by floristic as well as structural/physiognomic features. More recently, computer simulations have led to a better understanding of the dynamics of global vegetation models, where most recently they have been used to predict species range shifts under climate change. For simplicity’s sake, often both climate and vegetation models are combined.
This requires a reduction of the parameters—such as plant cover, water balance, biomass, soil carbon and many more—chosen for a single vegetation model. Such a reduction with respect to vegetation has even been proposed using only two plant functional types (Brovkin et al. 1997). At least two problems arise: there are too many parameters and the choice of parameters may always be comprehensible but still be arbitrary. Modellers are thus faced with the challenge of answering many questions: Should plant species, plant growth forms or plant functional types be taken into account? Further, what scale, grid size and time steps should be used for data collection in the field? The attempt to simplify often complicates one’s task because major uncertainties still surround the question of what variables to include. A model based on all ecologically important factors collected from the field for landscape and global spatial scales can prove troublesome when the focus is providing actual vegetation cover. Developing models that can accurately predict reliable changes for the future vegetation dynamics is extremely difficult and represents a major challenge facing the scientific community today.
However, recent developments in global ecosystem modelling have led to much more detailed representations of the world’s vegetation types, for example, the Lund-Potsdam-Jena (LPJ) Dynamic Global Vegetation Model. Such models are based mainly on land surface biophysics and on plant functional types with different physiological, morphological and phenological attributes with field data evaluation on different time steps (Sect. 22.1).
Equilibrium Theory of Island Biogeography.Relations between the distribution and establishment of organisms and the size of areas have been analysed in particular for islands. Islands are clearly bound with a limited number of different habitats, often under relatively uniform climatic conditions. They occur in various sizes and are situated at different distances from the mainland. Therefore, they have been chosen as examples to clarify basic relations between number of species and size of area.
Two main questions must be answered:
1. How do plants establish themselves on islands?
2. What limits the number of species on islands?
The first question is easily answered. Dispersal to islands situated far from the mainland, and thus from the closest source of propa- gules, can only be achieved by a form of long-distance dispersal. Transport by sea birds may be a method of dispersal (particularly epi- zoochoric), but also abiotic vectors, such as air and sea currents, or exceptional events, for example, tropical cyclones, must be considered.
On islands in closer proximity to the mainland, birds remain important; however, at greater distances dispersal by wind and water is more prevalent. Propagules can be transported to islands from several directions. Almost half of all species on Hawaii originate from the Indo-Pacific region, more than a quarter from the Holarctic and 17% from cold temperate Southern Hemisphere areas (Fenner 1985). A simple equation was developed to explain how plants can become established on islands. Arrhenius (1921) showed that the relationship between the size of an island and the number of species it maintains can be described by

Or
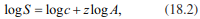
where S is the number of species of a taxon on the island, A denotes the area, z is a constant that changes little worldwide (i.e. the slope of the linear regression, when log S is plotted against log A, has values between 0.17 and 0.4), and c is a constant of proportionality and is dependent on the dimensions in which A was measured, in terms of the biogeographical area and taxonomic group.
The relationship between species and area is shown in Eqs. 18.1 and 18.2, where the number of species of a given group is halved if the area is reduced by a factor of 10. This attempt to provide a mathematically comprehensible theoretical expression relating the number of species to the size of islands is one of the important bases of the equilibrium theory of island biogeography by MacArthur and Wilson (1967). This has been confirmed, for instance, for plant species of the Scottish islands (Fig. 18.10). These relationships are explained by two hypotheses: (1) the habitat diversity hypothesis (Gorman 1979), which suggests that on larger islands there is a greater number of diverse habitats, and (2) the area-alone hypothesis (Kohn and Walsh 1994), which assumes a direct relationship between island size and number of species.
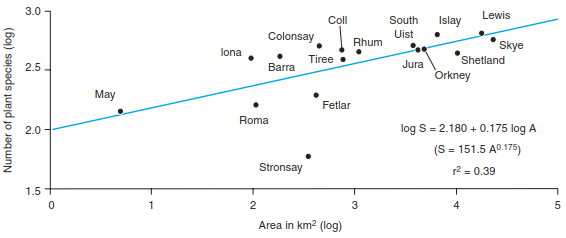
Fig. 18.10. Species-area relationships for plant species from selected Scottish islands (after Johnson and Simberloff 1974)
MacArthur and Wilson (1967) built on previous knowledge about the establishment and number of species on islands. They started from the following premises:
- Fewer species occur on islands than in the same area on the mainland.
- The number of species on islands increases exponentially with the size of the island.
- The pool of species on the mainland supplies propagules.
- The distance from the mainland is a very important determinant of the number of species on an island.
In addition, the researchers assumed that the number of species on islands depends on the diversity of habitat, and that if other islands exist between the island and the mainland, those will play an important role as a stepping stone. It was also concluded that islands possess a limited capacity for harbouring new species, which results in a sensitive equilibrium between the rate of colonisation and extinction. It should be noted that some islands may be “oceanic” islands (e.g. formed by volcanic eruptions) and as a result have never had direct contact with the mainland, or “continental” islands, where species contact has existed as a proportion of established species originating from some period of contact.
In summary, these observations and considerations lead to the formulation of a general theory attempting to explain the distributions of organisms found in island ecosystems. Ultimately, the most important parameters for such an explanation are the size of the island and its distance to the mainland. As such, three general conclusions were drawn:
- Larger islands have more species, also known as the “target effect” (higher colonisation rates on larger islands because they represent a larger “target”).
- Increased distance from the mainland (supplier of propagules) results in decreased numbers of species.
- At a constant number of species there is a continuous turnover of species due to colonisation and extinction.
The course of colonisation and extinction is shown schematically in Fig. 18.11a, where the intersection of both curves shows the equilibrium state, where similar rates of colonisation and extinction lead to a constant number of species, that is, the loss of species is compensated by the arrival of new species. While the number of species at equilibrium is constant, the identity of species and species composition may change, that is, there is a turnover of species.
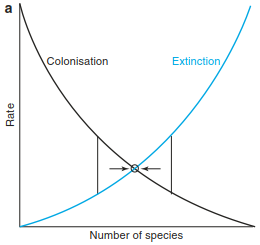
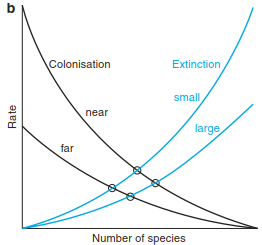
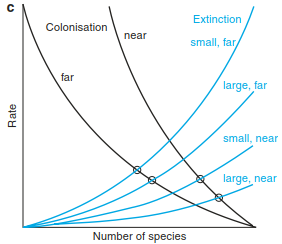
Fig. 18.11. Basic concepts in the theory of island biogeography. a Processes of colonisation and extinction. b Equilibrium (after MacArthur and Wilson 1967). c “Rescue effect” according to the equilibrium theory (after Brown and Gibson 1983)
The rate of colonisation (establishment on the island) is dependent on the distance from the mainland. The greater the distance, the more difficult it is for a species to become established there. The quality, type and quantity of the “source” of prop- agules that might arrive on the island must also be noted. The rate of extinction, however, is determined by the size of the island and will be much higher on small islands than on large ones because small islands can only support smaller population sizes, which have a higher risk of extinction. These conclusions have led to additional modification (Fig. 18.11b), where the following points may be deduced:
- Small islands have smaller numbers of species and higher turnover rates than large islands.
- Islands near the mainland have more species and a higher turnover rate than those further away.
- An island near the mainland returns to equilibrium after disturbance more quickly than one further away.
The equilibrium theory (or steady-state theory) was not generally well accepted at first. Some of the predictions of the model were verified empirically by Simberloff and Wilson (1969), who observed four islands off the coast of Florida following complete defaunation (sterilisation) and found re-establishment with the expected number of species and turnover rates in relation to size and position of the islands. Bush and Whittaker (1991) reconstructed rates of colonisation and extinction for spermatophytes on the volcano Rakata (Krakatau islands) on the basis of expedition reports and confirmed this theory as well (Fig. 18.12). On the other hand, different observations suggested that the extinction rate was dependent not only on the size of the island but also on the position of the island relative to the pool of propagules. The turnover of species on islands near the pool of propagules is rather low because of the continuous supply of propagules, which is of greater significance than the size of the island, which is known as the rescue effect, that is, a decreased rate of extinction due to recolonisation and immigration (Fig. 18.11c).
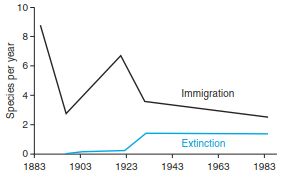
Fig. 18.12. Immigration and extinction of vascular plants on volcano Rakata, Krakatau islands, between 1883 and 1989 (after Bush and Whittaker 1991)
Criticism of the MacArthur and Wilson (1967) model is also directed at the type of predictive mathematical models used. Barkman (1990) believed that to describe complex functional relations in ecosystems, only descriptive models with limited validity should be considered. The characteristics of individual plant species and their interactions among each other have not been taken into account at all in these models. It is also assumed that the increased number of species is exclusive due to colonisation; genetic evolution is not considered, nor are the aspects of species saturation.
Lomolino and Weiser (2001) suggested that species richness may vary independently of island area, especially on small islands, and they termed this the small island effect. Responsible for a higher and more predictable speciation rate on larger islands is the internal geographical isolation of very different habitats such as large river basins and high mountain ranges, which are necessary for in situ speciation. Within-island spe- ciation rates can exceed immigration as a source of species richness, at least on islands larger than 3000 km2.
It has also been argued that phylogenetic diversification takes place on the same time scale as immigration and extinction. In this case, species richness on islands seems to be almost independent of all discussed factors. Finally, in a number of recent publications it has been highlighted more frequently that habitat heterogeneity, spatial scale and time scales (geological) are not sufficiently taken into consideration. Whittaker et al. (2008) proposed a general dynamic model (GDM) that provides explanations of biodiversity patterns by describing the relations between speciation, immigration and extinction, taking into account the life cycle of islands because climatic changes and geological tectonic events question the equilibrium model as well. In a proposed GDM, the most important variables considered remain—speciation, immigration and extinction—but are now combined with the evolutionary history of single islands and complete island archipelagos. The age of islands is also now taken into account; in particular, the geological history and the major tectonic events help to extend the thus far discussed conceptions of the equilibrium theory of island biogeography.
The theory of island biogeography also demonstrates that ecological equilibrium does not imply that ecosystems are constant and unchanging. The theory is based on a stochastic dynamic equilibrium, with a constant change in the rates of colonisation and extinction, which results in interannual variability for the actual number of species on islands and does not include specia- tion. Today it is becoming even more difficult to test empirically the simple assumptions of the theory of island biogeography simply due to the increasing anthropogenic influences. Although this theory has been criticised, it will be increasingly difficult to dispute, as empirical studies become increasingly harder to conduct, where the theory’s application can go beyond oceanic islands.
"Oceanic"and "Mainland" Islands
The results of island biogeography were formulated for islands and island groups surrounded by seawater and thus lack application for terrestrial island-like habitats. Nevertheless, attempts have been made to transfer this knowledge to the mainland and island-like habitats. These habitats are islands in lakes, mountain peaks in mountainous areas, “Inselberge” in the tropics and very small, well-isolated systems such as individual deciduous trees in a coniferous forest, caves or flower heads. Nowadays, all anthropogenic forests or biotope fragments in our managed landscapes could be interpreted to some extent as islands.
However, there are a number of commonalities to justify the application of island biogeography models to mainland islands. They have defined areas, relatively sharp borders with neighbouring habitats, are smaller than the surrounding area, and are often situated in a hostile surrounding area, at least for the taxa occurring within them. However, differences should not be overlooked, such as isolations (genetic separation) and separations (spatial separation). Usually, the distance to neighbouring islands (e.g. other forest fragments) is not very large, and the surrounding area may be hostile, but it allows at least some shortterm bridging. Most of all, species turnover is faster because immigration rates are higher (e.g. from species avoiding intensive agriculture and fleeing into residual forests), as are emigration rates (e.g. because of the relative proximity of comparable neighbouring islands or sudden external disturbances).
The disappearance of a species in a habitat does not necessarily result in the extinction of the species at all. If the tree line in mountains gets lower because of a climate cooling, species will probably find refuge in the valley. Ultimately, the risk of extinction is low. With subsequent warming certain species will become established again in higher altitudes. The same principle can be applied to the diversity of species in regions that became mainland species during prolonged cold periods and then, owing to certain climatic constraints within the species, became islands again in warm periods. In this case the equilibrium theory was contrasted with the relict theory, which states that the present-day occurrence of species and communities is the result of changes in the past.
For intensively used landscapes with many small habitats a mosaic concept was developed (Duelli 1993). The number of species is explained as individual “stones” in a mosaic; within the habitat the number of species increases with the number of habitat types (even those created by humans). In a mosaic landscape there are several transitional stages (ecotones) that may be colonised by specialists. Among animals habitat diversity favours those that are dependent on a seasonal change of habitats. This concept underlines the importance of transitions and edges between neighbouring habitats.
For small forest islands, species-rich edge zones can be observed where light-demanding species occur, but not typical forest species. In near-natural ecosystems, edge zones often act as buffer zones that reduce the immigration of external species to the inside of the island, which provides the habitat for obligate forest species. In intensively used agricultural landscapes such buffer zones are often lacking and the environmental gradients to the island edges are steeper. The relationship between area and number of species may thus differ according to the proportion of the edge and the core zone. In small forest patches species composition is determined by the species of the edge zone. With increasing area, species density decreases and species typical of continuous forest zones appear. The highest diversity is achieved within this transition zone between the edge and the core. In special cases this apparently simple relation is complicated by the differences in the quality and range of influences within this zone. A distinction must also be made between natural (wind, radiation) and anthropogenic (emissions, fertilisers, mechanical disturbances) environmental factors.
The function of habitat fragment corridors or stepping stones between larger mainland islands should also be noted. These may stimulate connectivity between habitats by assisting movement across the landscape, that is, by allowing transient residence of taxa without providing a permanent habitat. At the same time they may act as refuges. It is assumed that species are able to be more successful in expanding their range when stepping stones are present. This has been empirically shown for the assisted movement of animal species in fragmented landscapes, for example, for birds (Fischer and Lindenmayer 2002), but evidence on plants remains unclear.
Plants must move within landscapes as vegetative parts (ramets), pollen or seeds, assisted by wind, water or animals (Sect. 18.2.1). Therefore, the characteristics of the spatial pattern within the mainland might be equally or even more important than that of the stepping stones or corridors. However, because plants are sessile organisms, they are much more limited in their ability to decide whether a habitat is “suitable” or “hostile”, where mobile species can respond more readily to gradients of resource availability. Thus, more recently, the simple corridor-matrix model of metapopulations and landscape ecology has been replaced by an integrative functional mosaic model, where the landscape is composed of patches of different movement and flow characteristics, which provides a much more natural interpretation of the relationships (Murphy and Lovett-Doust 2004).
There are examples showing that mainland islands serve as solid proof of the validity of species-area relationships according to the theory of island biogeography if the influences of humans on the island are not too significant (Fig. 18.13a). These were confirmed, for example, for island mountains on the Ivory Coast, where the number of species increases steadily with increasing area (Porembski et al. 1995). However, in the tropical rainforests of Malaysia, increasing areas led to a significant saturation of species (in the reported case with an area of around 50 ha) (Fig. 18.13b). This contradiction has far-reaching consequences, particularly since nowadays extinction rates of plant and animal species are on the rise. For example, owing to the increasing losses of tropical rainforests, species extinction rates are calculated on the basis of species-area curves. The course of the curves in the figures shown here therefore allows a (too) broad spread of extinction rates. The saturation rate in tropical rainforests for an area of about 50 ha also indicates that plant sociological research examining minimum areas is possible theoretically but not practically.
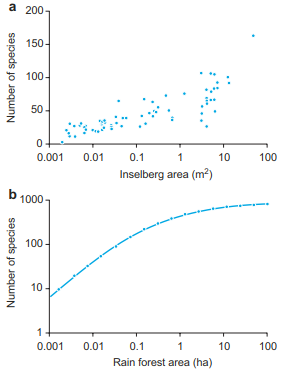
Fig. 18.13. Relationship between the number of species and habitat area. a On “Inselbergs” on the Ivory Coast (after Porembski and Barthlott 1993); b In a tropical rain-forest in Malaysia (after May and Stumpf 2000)
Because most of us today live in managed landscapes that typically have a high degree of fragmentation, it becomes more relevant to apply the theory of island biogeography to current problems, most prominently for questions surrounding protected areas. The relation of core to edge zones in small habitats becomes extremely important. With a spatial decrease in such habitats the diversity of conditions at a site decreases linearly, but the quality decreases exponentially (Mader 1983). Species that are locally adapted to specific site conditions may go locally extinct if abrupt changes in these environments occur. In contrast, more widely distributed species, which have broader habitat requirements, may suffer limited consequences in these edge environments, unless habitat is lost due to a large or frequent disturbance. Therefore, application of the theory of island biogeography is, at the moment, only limited. Akatov (2012) discusses in more detail the shortcomings of this theory, its possible uses and recommendations for practical planning of nature conservation projects.
In spite of these known shortcomings, conservation planning is still mainly based on island theory, although established relations between the size of an area and distance (for mainland islands, the distance to the next suitable habitat) have stimulated further discussions on the minimum size of areas that should be sustained. Of course, the size of a protected area should not be calculated according to the assumptions of island biogeography exclusively. Habitats of the same or similar quality should be maintained not too far away as (perhaps only intermittently required) refuge areas, in connection with the previously mentioned stepping stones. These stepping stones are an important component of the concept of biotope connectivity for protected areas. In our agriculturally managed landscapes, such stepping stones could be small forest islands (refuge) as well as hedges, edges of fields, long-term fallow land, abandoned stone quarries or railway lines. Concepts of nature protection must incorporate, along with the protected areas, buffer zones and smaller areas of habitat fragments around the protected area, and these areas should be maximised.
Preserving a given number of species should not be the sole aim of protection measures. Large areas of mosaics of optimal and suboptimal habitats should be the primary goal of such an area. As such, there is no one-size-fits-all approach when considering the size and spatial patterns for a protected area and stepping stones (particularly the distances between stepping stones and the protected area). Each of these aspects depends on the communities one is interested in protecting and can vary substantially. Very few empirical studies have been conducted to determine the minimum distances and areas that should be used for individual groups and organisms. Currently, urgent attempts to create protected areas are based on local conditions, the plausibility of plans and the availability of land. Increasingly, the more dynamic metapopulation concept and especially the functional mosaic model are becoming more important to help determine these issues of protected areas. The latter not only considers the dichotomy between unsuitable and suitable habitat patches but underlines the nature of the composite landscape mosaic as a key determinant of the fate of plant populations.
Date added: 2026-04-26; views: 155;