More Models of Island Biogeography Related to the Number of Species and Area
The theory of island biogeography was taken as a paradigm. Since the 1970s and 1980s various alternative interpretations have been advanced and discussed. Those who emphasise stochastic processes explain the establishment of plant species on islands differently to those who favour determinism. All theoretical considerations are based more or less on the same factors, which are regarded as decisive for the establishment of plants; however, they are interpreted differently. As the state of knowledge currently stands, the following factors have been identified as important in this theory: the pool of propagules on the mainland, mechanisms for dispersal, distance to the island and its size, habitat characteristics, phases of succession and the conditions of competition on the island.
Connor and Simberloff (1979) assumed in their simulation model that the establishment of plants occurs stochastically without competition. They tested this model with actual data and concluded that competition does not play a role. In their interpretation, it was the parameters of dispersal that determined the success of a coloniser. Gilpin and Diamond (1982), regarding competition as the decisive factor, criticised this interpretation. They mentioned that closely neighbouring islands possess different species compositions, although an exchange of species would be possible. However, this exchange does not take place because the appropriate species niches are already occupied, where invading species will not displace species already established. In this more empirical model the explanation of diversity on islands is not possible without considering competition, so the species first present will be a contributing factor to succession and the plant community.
These and other models use only those parameters that are considered important. Often, the dimension of the temporal scale has not been considered. Time has been shown to be an important component for the establishment of different species, as pointed out by the models developed by Grime (1979) and Tilman (1988, Sect. 17.3). For the early establishment phase, a stochastic interpretation is more relevant, whereas for the later phases, a deterministic one is possible.
Models developed by Whittaker and Jones (1994), which criticise purely stochastic models, show how much the immigration of species depends on the dispersing vector and on the stage of succession, as exemplified by their data from the island of Krakatau during the course of colonisation over 100 years (Fig. 18.14). They found that during the initial phase, colonising species reaching the island became established on undeveloped substrates. During the second phase, anemochoric species became more important.
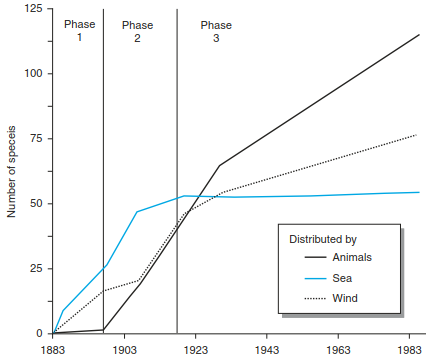
Fig. 18.14. Pioneer spermatophytes on volcano Rakata, Krakatau islands, from 1883 to 1989. The calculation is based on the assumption of minimum turnover. On the y-axis the data for number of species observed during different expeditions are shown. Species introduced by man are not considered (after Whittaker and Jones 1994)
However, it was only during the third phase, after substrates and the first succession stages of vegetation had developed, that zoochory played an increasingly important role. These results also suggest an initial stochastic but later deterministic establishment of species. It is overlooked in many interpretations that the turnover of species is not an inherent feature of a system but is largely regulated by changes in the environment. In this analysis and calculations, a minimum turnover rate is considered, but actual speciation rates are not available.
Date added: 2026-04-26; views: 157;