Plant Dispersal. Traits and Vectors
Introduction.The previous chapter showed how plants and plant communities develop over time and focused on short- and long-term temporal dynamics. These temporal dynamics lead to varying distributional patterns of species and communities. It is the task of spatial ecology to recognise such distribution patterns, to describe them, and to mechanistically explain their formation. Recognition and understanding of both the temporal and spatial dynamics of plant communities are indispensable for a thorough comprehension of biotic interactions within ecosystems, which is the topic of discussion in this chapter. The current chapter deals with the ecological basis of plant distribution, followed by a discussion of the relationships between species and area, with a particular emphasis on island biogeography.
Plant Dispersal.The life cycle of sessile plants includes a mobile phase, propagule dispersal. Plants will typically use one of the following two strategies: production of (1) generative propagules, where the plant produces spores, seeds, fruits, syncarps or even complete plants carrying seeds, and of (2) vegetative propagules, which include bulbs, shoots, runners, adventitious buds, parts of rhizomes and clonal growth (Fig. 18.1). Depending on the strategy of the plant in question, the distance travelled by the propagule will vary significantly and can be influenced through the help of abiotic or biotic factors. The aim of this process is to find a site that will facilitate the establishment and long-term survival of the plant, a so-called safe site. Finding a suitable site on which to become established is an extremely important stage in any plant’s life cycle, particularly in a highly competitive site. In any case, dispersal is crucial to the future presence of the plant’s population and to the plant’s capacity to persist in the vegetative community.



Fig. 18.1. Plants have many different methods of effective dispersal. a Aeloropus littoralis is a salt-tolerant coastal grass which spreads by vegetative runners (blastochory) as in south-western Arabia. b Self-dispersal (autochory) in Trifolium stellatum takes a special form (herpochory) with awns on the fruit having hygroscopic characteristics which enables the seed to move over short distances. c A special form of secondary animal dispersal (zoochory) is provided by coprochory. Dung beetles (many types of scarabs) collect the droppings of animals (e.g. cattle) and form it into balls. The dung contains seeds that are then buried just below the soil surface, where they may have a chance to germinate (Photos: K. Müller-Hohenstein)
Successful plant dispersal is typically improved when propagules possess certain morphological traits that improve their likelihood of finding an ideal site on which to become established. These traits can take advantage of external vectors to facilitate transport, which could include wind, water or even animals.
Traits and Vectors.Propagules are the functional units of plants and have adapted over time to be transported by wind, water, animals or through their own means of dispersal. Such traits include, for example, morphological structures on seeds, such as barbs and hooks, which cling to fur or feathers and can be transported long distances by the animal it gets attached to. Lightweight seeds or those possessing samaras (wings) can have significantly increased dispersal distances from the parent plants through wind. Nutritious, fleshy fruits attract animals by their smell, taste and colour. Most of the characteristics of seeds connected with transport are special parts of the pericarp. Some seeds still have adequate resources (coupled with resistant seed coats) for germination even if they have been eaten by animals, and they eventually find an ideal substrate medium (e.g. seeds from Argania spi- nosa, which are eaten by goats). Some dispersal strategies, modification and derivation are outlined in Table 18.1. Relationships between propagules and dispersal agents, especially by vertebrates, were compiled by Howe and Westley (1986) (Table 18.2).
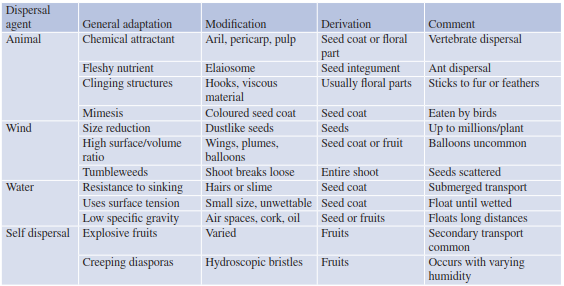
Table 18.1. Major dispersal strategies of fruits and seeds (after Howe and Smallwood 1982)
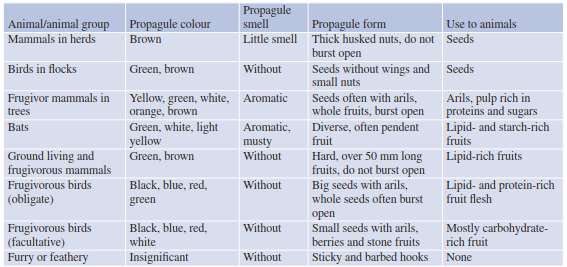
Table 18.2. Relationship between propagules and dispersal by vertebrates (after Howe and Westley 1986, with additions)
Dispersal vectors may change over time depending on environmental conditions. Species that can use multiple dispersal vectors are considered “polychor”. Many plants do not use only one vector but possess different morphological adaptations to exploit several vectors. The ability to use several strategies improves a plant’s chances of finding an ideal site for establishment. In special cases, however, a “secondary” dispersal strategy could prove more important. For example, an African elephant eats its favourite fruit, containing seeds, which are passed later in the day, 20 km from where they were eaten (primary dispersal). The surviving seeds are then further dispersed by dung beetles, which transport the seeds over short distances before burying them (secondary dispersal) (Fig. 18.1c) (Engel 2000). The importance of “secondary” dispersal is often underestimated. The example of dung beetles shows that the most favourable microsites are often only found in the secondary transport phase. Secondary transport may also lead to a wider range of dispersal.
However, the great variety of plant traits and different means of propagule transport requires a structured overview of the vectors and mechanisms associated with dispersal. Two large groups and many small subgroups are distinguished:
1. Propagation by the plant itself (autochory)
2. Propagation by other vectors (allochory)
Autochory.Over time, plants have evolved various mechanisms to ensure successful dispersal. In its simplest form, autochory, propagules are dispersed via gravity. Typically, the highest proportion of seeds is found closest to the parent tree, which is also known as the seed shadow. On rare occasions (i.e. steep slope or following a disturbance), dispersal can occur over longer distances. Such plants have no special traits for dispersal.
Propagules can also reach suitable growing sites by growth processes, for example, via scions (vegetative shoots) or flower stalks (pedicels, e.g. Cymbalaria muralis). Propagules are even able to move by themselves over short distances, for example, by twisting awns due to changes in humidity (Fig. 18.1b). In addition, some plants have even evolved mechanisms to project propa- gules by a single impulse or shot, which helps ensure further distances travelled from the parent plant. The propagule ejections can be triggered by a series of different vectors: animals, wind or even a simple raindrop (e.g. Impatiens noli- tangere). In some species, these triggers are not required, where certain changes in abiotic factors must occur for projection of the propagule to occur. For example, differences in turgor pressure in a plant (e.g. Ecballium elaterium) or changes in moisture exchange through the seed coat (e.g. Bauhinia purpurea) can trigger this phenomenon. Seed dispersal through this mechanism can easily cover several metres in some cases.
Allochory.Wind is an important abiotic dispersal vector for allochoric seed transport (anemochory). Even when seeds reach the ground or soil substrate, they can continue to be blown along the surface. Plant propagules have been able to adapt morphologically to ground dispersal, especially in dry arid regions of the world where there are rarely impediments or barriers to such transport. More importantly, however, is transport by air currents. Small, light propa- gules are particularly benefactors of this mode of dispersal as they can easily be dispersed multiple times under the right gusts of wind, for example, seeds of orchids or spores of cryptograms. Some other interesting wind-dispersed adaptations include balloon fliers (e.g. Astragalus spinosus) that possess special morphological adaptations, as do seeds or fruits with parachutes (e.g. Taraxacum officinale). A number of tree species (e.g. Acer and Fraxinus spp.) have adapted special wings (or samaras) to help keep propagules in the air and increase dispersal distances. Wind also triggers seed transport in plants that scatter seeds (e.g. Papaver spp.).
Hydrochory involves the transportation of propagules either in or with the help of water as a dispersal vector. Some propagules are able to float because of special tissues or large intercellular spaces that have a low specific weight and that typically possess a hydrophobic seed coat, which makes it difficult for the seed to absorb water (e.g. Nymphea species). Coconuts and certain seeds and seedlings of some mangrove species are excellent examples of this adaptation, where they are able to stay in salt water for extended periods of time with little or no effect on seed viability. A seed that falls into a moving body of water (i.e. river, lake or ocean) can almost ensure successful long-distance dispersal; however, a body of water is not always required. In drier, more arid areas, seeds can travel substantial distances from their parent plant with the help of a few raindrops (e.g. Anastatica hierochuntica in dry regions). There are disadvantages to this mode of dispersal, however, as floating for prolonged periods of time can cause premature germination with a lack of an anchoring substrate. Plants usually prepare their offspring for this encounter by certain dormancy cues, where a seed must meet specific environmental conditions (typically moisture and temperature) prior to breaking dormancy and allowing germination to ensue.
The most important and ecologically most complex form of allochoric dispersal that exists is zoochory. Zoochory involves the use of animals as dispersal vectors for plants. Plants have developed different mechanisms and traits to entice or attract animals to disperse their seed, where, interestingly, animal foraging and behaviour can determine patterns of seed dispersal. Close interrelations between some plant and animal species point towards a long co-evolutionary development. This applies particularly to endo- zoochory, where propagules are transported internally by the animal. To do so successfully, seeds should have a relatively hard shell that ensures they are resistant to conditions within the animal’s digestive system (i.e. high acidity). After excretion, seeds are usually provided with good starting conditions for germination in the nutrient-rich excrement. Although propagules can be taken up randomly, very close links between birds, bats and herbivores with plant species through the use of fruit can entice attraction and increase the likelihood of seed dispersal. These are especially evident in the humid tropical regions of the world (Table 18.2). Zoochory does have some disadvantages as a dispersal medium; foraging animals can hoard or cache seeds (i.e. squirrels) in a safe site, which could reduce the likelihood of future germination (e.g. seeds of Pinus cembra collected by Nucifraga caryo- catactes), but some of them still have a chance to survive because not all hiding places are found again and some seeds are also lost during transport.
Birds (ornithochory), bats, ants (myrmeco- chory) and many larger animals are especially attracted to the fruit of plants. Plants invest in nutrient-rich fruits and have evolved to attract dispersal vectors by the use of bright vibrant colours and strong odours. Ants may carry seeds possessing lipid-rich elaiosomes (e.g. Corydalis cava), but the seeds remain untouched. Birds are especially attracted by the colouring of fruits and are even able to distinguish ripe seeds from unripe ones. Sometimes propagules are eaten or damaged. These are often starch-rich and can provide nourishment for an animal. In some instances a monkey, for example, will ingest the fruit pulp and then spit out the seed.
Epizoochory is a form of passive dispersal by animals, where the seed possesses mechanisms to cling to the hair or fur of the animal. Typical mechanisms include glue-like excretions, glandular hair, barbs with awns and other outgrowths formed from the pericarp that can attach to the animal. By doing so, the seed greatly improves its dispersal distance; however, unlike in earlier cases, the animal is not rewarded for its transportation through nutrient supplementation. In endo- zoochory, close interaction exists between plant and animal, where dispersal is regulated, sometimes even targeted. As a result, epizoochory is mainly random.
Two additional dispersal mechanisms exist that do not really fit well with the previously mentioned groups. The first is atelochory (also called achory), where dispersal does not occur, and in fact it is prevented. The consequence of this evolutionary development is that reproduction takes place at the site where the mother plant grows, which is favourable to the species. This lack of dispersal allows the offspring to take advantage of similar conditions its parents have.
Arachis hypogaea or Trifolium subterraneum provide examples of such a strategy, where, following pollination, the pedicel and ovary penetrate into the soil and become established.
The final mode of dispersal for propagules involves humans (anthropochory), which with the onset of the Anthropocene has played an increasingly important role in the recent history of plant dispersal. In this particular form, any dispersal distances can be achieved, and essentially, no geographical or ecological barriers exist. Anthropochory dispersal occurs when humans are looking to move certain species of plants to specific areas for either food or ornamental purposes. If dispersal occurs unintentionally along with other propagules (e.g. weeds disperse along with crops of cereal), their distribution and the pattern of the species concerned typically almost always occur randomly. Both forms are closely linked to the problems with neophytes (Sect. 17.3).
Date added: 2026-04-26; views: 157;