Vegetation Dynamics and Strategy Models
Variability within a single plant can be considerable, growth can be fast or slow, tolerance to shade can be high or low, and dispersal from the parent plant can be far or near, to name a few parameters. Such characteristics, acquired during the course of evolution, enable plants to successfully compete with other individuals and to become established and are often summarised by the term “strategy”. It is defined as the sum of genetically fixed physiological and morphological adaptations required to cope with different environmental conditions, conquer a habitat and persist with optimal use of resources. Individual plants, populations and species are also classified using strategic aspects as plant functional types (Sect. 20.2).
McCook (1994) showed how two plant species with different characteristics may behave in competitive ways (Fig. 17.41). The following basic variants are possible. One species gets suppressed at different temporal rates, both species achieve a similar abundance, or both species become dominant at different times. All strategic models are based on such assumed forms of performance and differ only in the way they are interpreted. External influences are not always considered (e.g. climate change, herbivory). Because overlapping anthropogenic interference is also excluded, the knowledge gained is relatively limited. Other models originate from functional characteristics of the whole ecosystem and ask which of these have changed over time and how. These models usually generalise too much, where some are exclusively descriptive, while others try to make predictions or attempt to explain current dynamic processes. Ultimately, no single model provides a comprehensive explanation for the empirical findings from the field, at least none has to date.
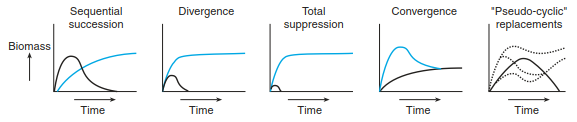
Fig. 17.41. Possible behaviour of two species over the course of succession. (after McCook 1994)
Initial FloristicComposition.The model of initial floristic composition (IFC), based on Egler (1954), is still used to describe the sequence of successions because it represents sequences that are physiognomically recognisable. According to this model, the complete set of species is present from the beginning, and a sequence of starts results in only a few of these species becoming dominant. Figure 17.42 shows the course of growth for six species from germination via establishment of seedlings, through to the vegetative stage, and finally to the reproductive growth phase. Species 1-4 are relatively short-lived therophytes to short-lived woody species, and species 5 and 6 are long-lived, shade- tolerant species. When the latter species are fully developed, the short-lived ones are no longer able to persist. IFC is still accepted as a descriptive model of primary as well as of secondary successions. There are no generally valid succession sequences because of external disturbances and stress situations caused by them. Egler introduced this concept of gradual change as the centre point and prefers the term “development of vegetation” to that of succession. The limitation of such a model is the exclusion of the propagule influx.
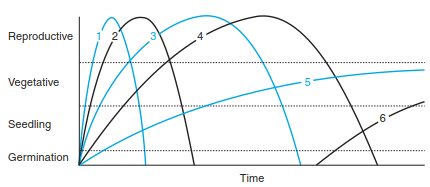
Fig. 17.42. Scheme of initial floristic composition model of succession. The curves show the behaviour of six different species. (after Finegan 1984)
r and K Strategies.The concept of r and K selection is based on models of population growth in animals, which was developed by MacArthur and Wilson (1967). This concept is used by botanists and zoologists to stress particularly contrasting characteristics of species. “r” is the intrinsic rate of population growth or rate of reproduction, while “K” represents the carrying capacity or maximum population size. Essentially, the r and K concepts stress the allocation patterns of limiting resources that are inversely related.
r-strategists form a large number of very small seeds representing a high proportion of their phytomass; they also tend to have an efficient mechanism for dispersal and grow fast without high biomass accumulation. This leads to a much quicker time to reach reproductive maturity, and such plants also tolerate long periods of seed dormancy, but the seeds can be activated very quickly. Species that are r-strategists are short-lived opportunists that are able to become established faster in their environment than shade-tolerant species, but they are inferior competitors to k-strategists.
k-strategists are long-lived and slow-growing and accumulate large amounts of biomass over time. For this reason, and because they are shade tolerant, they are able to outcompete r-strategists.
They invest less into reproduction and will typically produce a small number of seeds that are larger than those of r-strategists. However, they need to form metabolites for defence to protect long-lived leaves and other organs from herbiv- ory. In terms of successional stages, it is easy to determine that early stages are dominated by r-strategists, while later stages are dominated by K-strategists. On fallow land, for example, annual and biennial species are replaced over time by shrubs, which are then taken over by trees.
The aforementioned characteristics are accepted as important trade-offs, but a continuum is seen between the extreme positions of r- and K-strategists. This continuum also indicates not only that disturbed habitats are colonised by r- strategists in the short-term (Fig. 17.43) but also that those habitats permanently dominated by K- strategists do not necessarily allow stress-free growth. With the requirement for stress tolerance, a further strategy type is indicated.
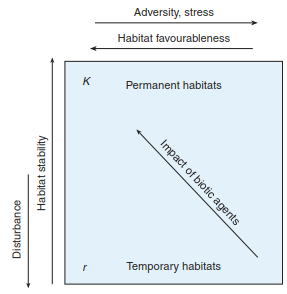
Fig. 17.43. Model of K- and r-selection considering effects of stress. (after Brown 1985 and Brown and Southwood 1987)
Strategy Types of Grime (C-S-R Model).Grime (1974) and Grime et al. (1988) worked out one of the most well-known models of vegetation dynamics: the triangle model of primary ecological strategies. Stress tolerance, adaptation to unfavourable conditions and reaction to disturbance missing in the r/K strategy model were regarded as particularly important. Three strategy types were introduced (Sect. 19.3):
Competitor strategist (C): long-lived, competitive species on favourable sites without limitation of resources and almost stress free. They use resources particularly well, often possess storage organs and show considerable plasticity in root and shoot formation, continuously produce leaves that live for only a short time, and have low seed production. These are perennial herbs, shrubs and trees, which form in mid to late successional stages.
Stress-Tolerant Strategist (S): species adapted to unfavourable sites with limited or poor availability of resources (e.g. inadequate light, drought, nutrient deficiency, frequent frosts). They are long-lived, but with low productivity and reproductive rates and are often restricted to sites with little or no competition. Extremely stress-tolerant species are lichens on cold and dry sites.
Ruderal Strategist (R): short-lived, usually herbaceous species with fast growth rates and high seed production, usually self-pollinated with rapid seed ripening and dispersal, disturbance-tolerant opportunists and pioneers, but weak competitors. Therefore, they grow on sites with frequent, natural disturbances.
However, it is difficult to classify many plant species into only one of these three strategies. Intermediates are frequently observed, where plants with characteristics of several strategic types in different combinations are possible. On favourable sites where competitor strategists should dominate, C-R transition forms are found if the favourable conditions are temporarily disturbed, for example, by mowing meadows. Stress- tolerant competitor strategists (C-S) are long-lived species; for example, many tree species of nemoral forests have adapted to non-optimum sites. Stress- tolerant ruderal strategists (S-R) grow on unfavourable sites that are often disturbed. Included is also an intermediate type (C-S-R) combining several strategies and may also occur in temporal sequence within the same species.
Grime et al. (1988) attributed life forms to some of his strategy types and included them in areas of his so-called strategic triangle (Fig. 17.44). These strategy types are linked to succession sequences, assuming different productivities. On productive sites, development proceeded from ruderals via competitively strong types to those plants with competitive-stress strategies. This applies in Central Europe to beech forests on limestone, where on unproductive sites the final stage is determined by stress- tolerant species. The more productive the site, the higher the proportion of competitively strong species in the mid-successional period. The intensity of competition increases with increasing productivity at the site. However, Tilman (1990) argues against this conclusion.
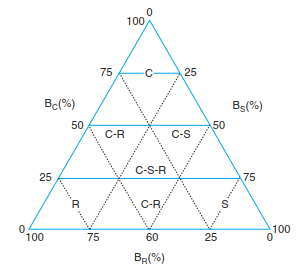
Fig. 17.44. C-S-R diagram of different types of strategy according to Grime. C competitor species; S stress-tolerant species; R ruderal and intermediate types with relative importance of competition (BC), stress (BS) and disturbance (BR). (after Grime et al. 1988)
Resource Ratio Model of Tilman.All plant species are limited in their distribution by resources, which are often difficult to acquire but are required for survival. A species is deemed successful in competition for such a limiting resource if it is able to reduce the level of nutrients to the lowest level (called R*), resulting in competitive exclusion (Sect. 19.3). Tilman (1982) developed this model to describe the growth of algae, while for terrestrial systems the assumption is made that the most important limiting resource is within the soil (typically nitrogen) or is light. Any limitation in either of these resources will lead to increased stress within the plant. Often, these resources are inversely related, that is, a low nitrogen supply is typically associated with favourable light conditions, and low light levels (in the understorey) are commonly found at sites with favourable conditions for plant nutrition. Gradients between those two extremes lead to succession stages where plants replace each other according to their demands and their specific competitiveness.
Each species grows fastest and reproduces best with the resources to which it is best adapted and where it can outcompete other species. Sufficient root biomass is very important for obtaining sufficient nutrients in the soil, but also sufficient above-ground biomass is essential for light capture by producing leaves. In primary succession, species able to cope with a poor nitrogen supply typically exclude species that require a great deal of nitrogen. As time passes, however, soil nutrition gradually improves thanks to mineralisation and nitrogen input into the system. Now, species able to cope with less light are more competitive. This hypothesis is the basis of the resource ratio model or the nutrient-light relation model.
Simpler than the ideas of this model are those of the nutrient-colonisation and light-colonisation models, or the “competition-colonisation tradeoff”. The former again applies to nutrient-limited habitats. Colonists producing many seeds, which they are able to disperse quickly, are initially successful. However, they are weak in the competition for nutrients, and thus species that allocate their resources in such a way as to establish their rooting system to gain more nutrients are favoured. The corresponding situation applies to nutrient-rich habitats for light as the limiting factor. These interactions for five grass species demonstrating their ability for competition and colonisation are shown in Fig. 17.45.
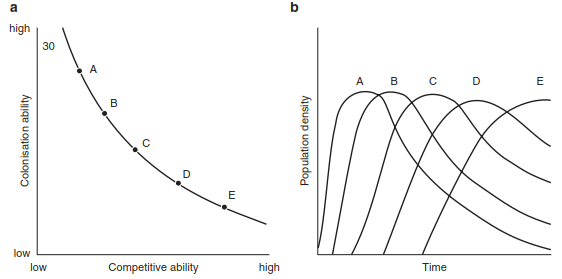
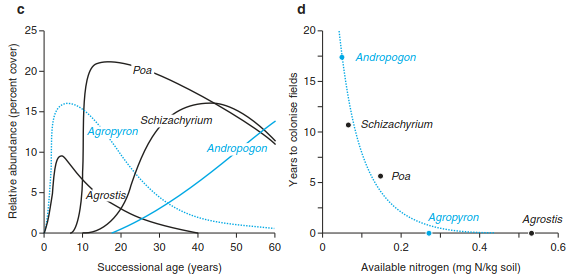
Fig. 17.45. Relationship between competition and colonisation capability. a Curve represents a hypothetical interspecific trade-off between competitive ability. Each point on this curve could represent a distinct plant species. The five points shown are for species A, B, C, D, and E. b Species A–E will have qualitative successional dynamics because of their colonisation–competition trade-offs. c Successional dynamics of dominant grass species of Cedar Creek Natural History Area (after Tilman 1988).
d Observed interdependence of competitive ability and colonisation rates for five dominant grasses of Cedar Creek Natural History Area. R* for nitrogen is the observed level to which monocultures of each species reduced the concentration of dissolved ammonium and nitrate on fertile, low-nitrogen soils (Tilman and Wedin 1991a). Lower R* values correspond to a greater competitive ability (Tilman and Wedin 1991b). The colonisation rate is inversely related to the years each species required to colonise newly abandoned fields at Cedar Creek Natural History Area. (after Tilman 1994)
From these models, the consequences for biodiversity within areas may be derived. The more heterogeneous a space to be colonised is—partic- ularly regarding its environmental conditions (e.g. nutrients, light, water) and natural and anthropo genic disturbances—the more species are able to coexist. It follows that with increasing heterogeneity not only does biodiversity increase but, in the habitat, resources should ideally be available at intermediate levels (Sect. 20.3).
Facilitation-Tolerance-Inhibition Models of Connell and Slatyer. Competition for resources may change by type and amount over time, especially when disturbance regimes are considered. Connell and Slatyer (1977) pointed out that the driving force behind vegetation dynamics and stress is the issue of scale, where certain dynamics only occur at certain scales. As such, three models, or succession pathways, are considered (Box 17.5):
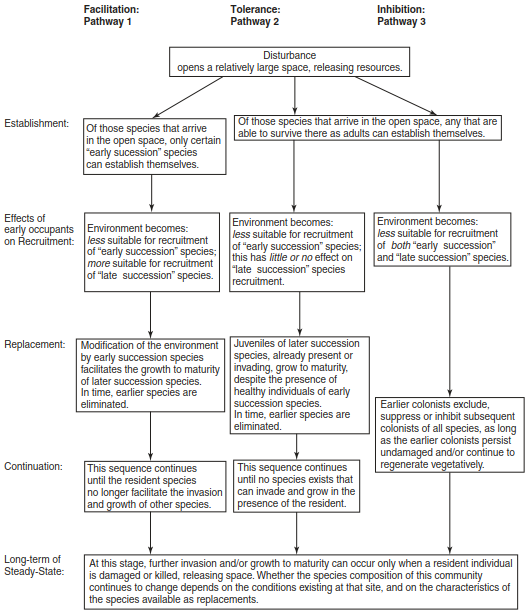
Box 17.5: Three “Succession Pathways” of Connell and Slatyer (1977) (after McCook 1994)
Facilitation model: Facilitation is understood as the “promotion” or “enabling” of other species, starting from the assumption that the first colonists change site conditions over the course of time autogenically in such a way (e.g. change in substrate, formation of humus) that colonisation by more demanding species is stimulated. This is usually linked to the displacement of the first colonists as they become unable to cope with the competitive advantage of the incoming species.
Tolerance model: Late successional species are neither stimulated nor inhibited. However, they are only able to establish themselves if previously established individuals die or are removed, leaving an area to invade. The species now present compete with one another and behave similarly to the resource ratio model of Tilman, where species with lower R* gain dominance.
Inhibition model: Earlier colonists suppress or inhibit subsequent invaders, which will only be able to become established in the event of a disturbance or by the creation of open space for short time spans due to an individual’s death and not because of their competitive advantage.
All three models have been tested in field experiments. The facilitation model may be observed in primary successions, where pioneer species prepare the substrate, as well as following disturbance, because the succession is thrown back into an earlier phase. Both of the other model versions were tested in later, species-rich successional stages. The final steady-state stage is regarded as relatively stable, but cyclical dynamics are not excluded.
Also, the question of stability is taken into consideration. Consequently, succession is regarded as a balancing reaction of a “stable” system that has been heavily disturbed. The spatial and temporal extent of the disturbance as well as their type and intensity are decisive for the following new initial stage of succession.
Date added: 2026-04-26; views: 158;