Vegetation Dynamics and Influence of Animals
At any time during succession, two important influences by animals must be taken into account (Sect. 19.4). The spectrum of species may be changed by herbivory, or at least the conditions for competition are changed. This is rarely fully taken into account because herbivores often eat seeds (granivores) and seedlings, and the extent of such disturbance is difficult to measure. Often it is a matter of speculation as to how vegetation would have developed without herbivory. However, animals are also vectors for propa- gules and, thus, for the distribution of species, and they participate decisively in the formation of spatial patterns and the composition of vegetation units. In addition, animals change site conditions by their excrement and by their mechanical activities and soil disturbance.
There is much evidence that animals, too, exhibit distinct dynamics. In the initial stages of succession, arthropod herbivores dominate, with 93% of all insect species; however, these proportions drop drastically within older successional stages (51%) and even more so in forest communities (44%). During the initial stages, sapsucking phytophages make up half of that population, later only a quarter. Macroinvertebrates and predators associated with them increase with the age of the succession.
Brown and Southwood (1987) tried to derive from these observations basic adaptation strategies for plants and animals during succession (Fig. 17.46). They concluded that faunistic partners in the early stages of succession are usually opportunists (r-strategists) investing their resources in their reproduction. Many insects are able to fly, promising fast colonisation. The phytophages are mostly generalists, and food webs are rather simple. In the mid-successional stage, secondary consumers are also represented among the animals, particularly insectivores. Food webs and the structural organisation of the habitat become more complex. Few of the occurring insects have wings, and the invertebrates are uniformly distributed. The high structural diversity of vegetation in the late successional phase begets a large number of niches for specialists. Morphological and trophic diversity in invertebrates (particularly insects) and vertebrates (particularly birds) is high, and ^-strategists dominate even for animals.
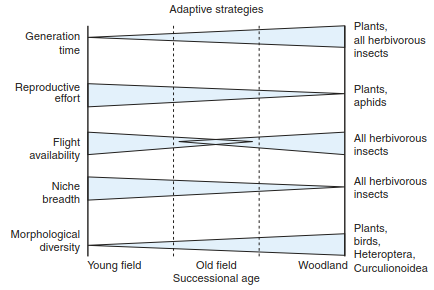
Fig. 17.46. Adaptive strategies of plants and animals during secondary succession. (after Brown and Southwood 1987)
Further Aspects for Understanding Vegetation Dynamics.In addition to the concepts of vegetation dynamics mentioned so far, other ideas that incorporate the enormous complexity of ecosystems could be discussed, for example, those concerning convergence and divergence in the development of successions (Leps and Rejmanek 1991), the continuum concept by Austin (1985) or the discussion of chaos theory and non-linear systems (Stone and Ezrati 1996). These models, as well as those with partially overlapping concepts covered previously, show that it is not possible to describe all aspects of vegetation dynamics with “one simple formula”. Complex processes and patterns of vegetation dynamics may only be generalised or modelled to a certain extent. Descriptive models of the sequence of individual species are accepted, but they explain little. Also, little emphasis is placed on the fact that a species performs differently in juvenile stages than in older stages. As summarised by Bazzaz (1990: 258), “Despite much progress in modelling succession, the tension between simplicity of models for prediction and complexity to include all relevant parameters remains unsolved.”
A path towards a better understanding of processes and mechanisms of vegetation dynamics should integrate previous and new information from experimental approaches. Note should be made of different spatial and temporal scales, as well as of the organisational hierarchies of plants—individual, species, population, community. Recently, dynamic simulation models have been used to describe vegetation dynamics. In southern Ecuador, Dislich et al. (2009) used the FORMIND model to simulate the dynamics of tropical mountain forests and regarded this model as being well adapted to investigate disturbances to forest structure and species composition. The empirical results of long-term studies should be used for verification. For example, the results obtained by Schreiber (1995) on very detailed succession studies over more than 20 years (on fallow land) do not fit into any succession model. This means that succession can hardly be exactly predicted, and planning based on such predictions is risky.
Date added: 2026-04-26; views: 152;