Ecological Stability of Plant Communities and Disturbances
The topics of succession and cyclical vegetation dynamics touch on questions of the “stability” of plant communities and whole ecosystems, as well as the concept of ecological equilibrium, sometimes naively termed the “balance of nature”. Much controversy continues to surround this complex topic, particularly in relation to the definitions of stability and of the characteristics required for stability (Grimm et al. 1992). Temporal and spatial scales are often not considered sufficiently: stability is related to momentary conditions, years or centuries, of plant communities considering a span of only a few square centimetres up to many square kilometres. In all cases, stability/instability is linked to species interactions and demographic stochas- ticity (internal sources) or disturbance, environmental change and stress (external sources). The relations between stability and diversity or complexity are discussed in Sect 20.4.
Generally, definitions of ecological stability are linked to a system capable of maintaining its structures and functions over the long-term and that is properly used by humans. Population sizes of species may fluctuate little around a mean value, and turnover of species is possible to a limited extent (but not that of life forms or functional groups). Functional relations, such as within food webs, and the many different niches and feedback mechanisms in a system strongly determine stability. Theoretically, stability may thus be explained via the sum of all functional connections within a system. It is a dynamic concept that also includes the stability of processes. Stable systems react to disturbance and return to the initial stage by self-regulation. Ecological stability includes cyclical changes, but not successions. In contrast, a labile system is not able to withstand changing external influences.
There is a clear differentiation between complexity and stability. For the complexity in a system, the number of species, connectance, interaction strength and the evenness of species distribution are most important. Several components of stability may be distinguished as additional variables of interest that could be applied to species abundances, species composition or ecosystem properties and processes within plant communities (Pimm 1984):
- Qualitative stability: property of a system that returns to its original state after a perturbation.
- Resistance: a measure of the ability of a system to maintain its original state in the face of an external disturbance or perturbation.
- Resilience: a measure of the speed at which a system returns to its original state after a perturbation.
- Robustness: a measure of the amount of perturbation that a system can tolerate before switching to another state.
- Variability: a phenomenological measure of the magnitude of temporal changes in a system property.
- Persistence: a measure of the ability of a system to maintain itself through time.
Disturbance and strain are generally seen as influences that indicate a “situation deviating from the norm” and that lead to stress situations. The extent to which a system can cope with strain depends on the type, intensity, duration and spatial extent of the disturbance (Fig. 17.47).
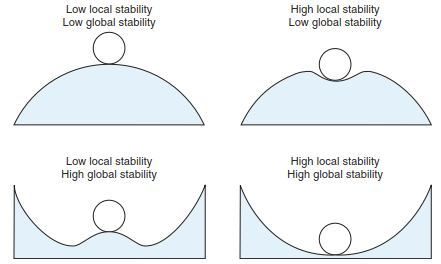
Fig. 17.47. Relations between different spatial scales (local, global) and stability. (after Begon et al. 1999)
Gigon (1982) also suggested that ecological stability should not only be related to some vaguely defined “balance”. Other stabilising mechanisms, such as inertia, buffer effects, the replacement principle and risk distribution, also affect ecosystems. Important characteristics are the basic dynamic behaviour of ecosystems and the occurrence (or absence) of disturbances. He also states that imbalances and instabilities in nature are just as decisive for evolution as balance and stability. Gigon thus distinguishes between the following aspects:
- Ecological stability, which includes the resistance of a system or its ability to return to its original state after a disturbance (resilience).
- Ecological instability, where changes after a disturbance cannot be reversed.
- Ecological lability, which is a transient phase with a strong disposition to change as a result of disturbance.
| Fig. 17.47 Relations between different spatial scales (local, global) and stability. (after Begon et al. 1999) |
High local stability Low global stability

|
| Low local stability Low global stability |
Forms of ecological stability may be described as constancy if no actual changes take place (except changes of individual plants) or as resistance if disturbances are buffered (Fig. 17.48). In these cases, the affected systems remain almost unchanged. Other forms where a major dynamic is apparent but the original state is still reached may be called cyclical with regular fluctuations and regeneration cycles or resilient if regeneration takes place after irregularly occurring disturbances.
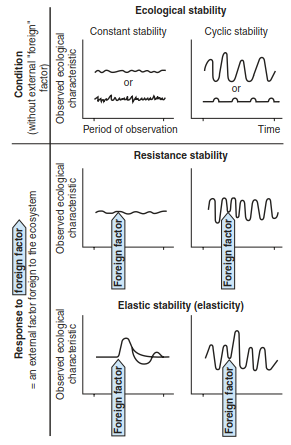
Fig. 17.48. Basic types of ecological stability. These types differ according to the dynamics of chosen characteristics and to the presence or absence of external factors. (after Gigon 1982)
Forms of instability (Fig. 17.49) lead to changes in the ecological characteristics of a system. These may be triggered endogenously (e.g. in autogenic successions) or exogenously (owing to disturbances). Instability may also result in endogenous or exogenous fluctuations (e.g. insect calamities or fluctuations of agricultural yield because of pathogen attack). These forms of stability and instability may occur in the same system. An individual tree performs differently when it is part of a complete forest; a locally destructive fire may be necessary for the maintenance of the system over a large area. Only a few cases fail to fit into this concept because of insufficient observations or insufficient knowledge of the interfering factors.
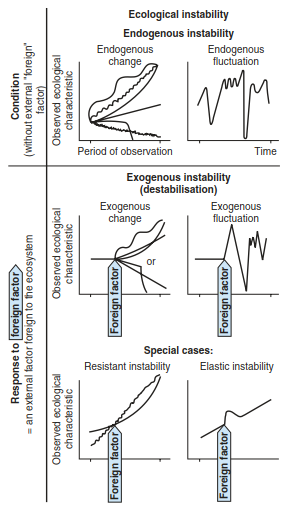
Fig. 17.49. Basic types of ecological instability. These types differ according to the dynamic of chosen characteristics and to the presence or absence of external foreign factors. (after Gigon 1982)
Disturbances or perturbations are not necessarily negative for the stability of ecosystems or communities. They can even be essential for the maintenance of a dynamic ecological equilibrium. A boreal larch forest would develop into quite a different system without the regular influence of fires. The same interfering factors may thus have different consequences for a system. Disturbances may be seen as inherent properties of a system or as stochastic external events, the latter caused naturally or anthropogenically. Several years without precipitation in desert regions is an internal disturbance to the system, as is the regulation of population density based on predator-prey relationships, or the dynamics of avalanches in montane ecosystems.
It is often difficult to distinguish between disturbances caused by endogenous biological processes and those caused by exogenous environmental influences. Variances in spatial and temporal scales must be taken into account. The scale extends from disturbances by continental drift, on a scale of millions of square kilometres taking millennia to occur, to damage by spring tides, late frosts or extreme herbivory affecting a few hectares over a much shorter time frame.
Finally, it must be stressed that there is no “balance of nature” in the sense of completely stable ecosystems: ecosystems are always dynamic and undergoing change. In addition, no value judgement is implied by the concept of ecological stability. A stable system might be judged differently by a farmer than by an environmentalist, by a skier differently than by a hunter. A practice-oriented synecology must include an evaluation of stability, which is based on accepted standards and models.
References:Austin MP (1985) Continuum concept, ordination methods and niche theory. Annu Rev Ecol Syst 16:39-61 Badal E, Bernabeu J, Vernet JL (1994) Vegetation changes and human action from the Neolithic to the Bronze Age (7000-4000 bp) in Alicante, Spain, based on charcoal analysis. Veg Hist Archaeobotany 3:155-166
Barnosky AD, Hadly EA, Bascompte J, Berlow EL, Brown JH, Fortelius M, Getz WM, Harte J, Hastings A, Marquet PA, Martinez ND, Mooers A, Roopnarine P, Vermeij G, Williams JW, Gillespie R, Kitzes J, Marshall C, Matzke N, Mindell DP, Revilla E, Smith AB (2012) Approaching a state shift in Earth’s biosphere. Nature 486:52-58
Bar-Yosef O (1998) The Natufian culture in the Levant, threshold to the origins of agriculture. Evol Anthropol 6:159-177
Bazzaz FA (1990) Plant-plant interactions in succes- sional environment. In: Grace JB, Tilman D (eds) Perspectives on plant competition. Academic Press, San Diego
Begon M, Harper JL, Townsend C (1999) Ecology, 3rd edn. Blackwell, Oxford Bick H (1993) Okologie. Fischer, Stuttgart Bohmer HJ, Richter M (1996) Regeneration—Versuch einer Typisierung und zonalen Ordnung. Geogr Rundsch 48(11):626-632
Brown VK (1985) Insect herbivores and plant succession. Oikos 44(1):17-22
Brown VK, Southwood TRE (1987) Secondary succession: patterns and strategies. In: Gray AJ, Crawley MJ, Edwards PJ (eds) Colonisation, succession and stability. Blackwell, Oxford
Date added: 2026-04-26; views: 168;