Resting Membrane Potential: Ionic Basis, Nernst Equation, and Goldman-Hodgkin-Katz Equation
At rest, neurons exhibit a potential difference across their plasma membranes, with the inside of the cell negatively charged relative to the outside (Figure 6.8). This electrical difference is termed the resting membrane potential (abbreviated Vm). By convention, the extracellular fluid serves as the voltage reference point, and the polarity of the membrane potential is stated as the sign of the excess charge on the inside of the cell. For example, if the intracellular fluid has an excess of negative charge and the potential difference magnitude is 70 mV, the membrane potential is reported as –70 mV (inside relative to outside). Importantly, volts measure the difference in charge across a membrane; a Vm of –70 mV does not indicate the absolute number of positive or negative charges on either side.
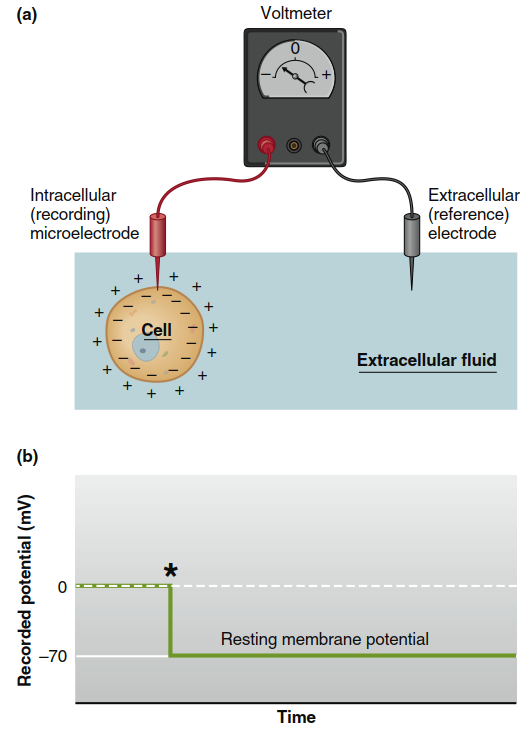
Figure 6.8. (a) Apparatus for measuring membrane potentials. The voltmeter records the difference between the intracellular and extracellular electrodes. (b) The potential difference across a plasma
membrane as measured by an intracellular microelectrode. The asterisk indicates the moment the electrode entered the cell
Nature and Magnitude of the Resting Membrane Potential. The magnitude of the resting membrane potential in neurons typically ranges from –40 mV to –90 mV. This potential remains steady unless changes in electrical current alter it; by definition, a cell under such conditions would no longer be “resting.” The resting membrane potential exists because of a tiny excess of negative ions inside the cell and an excess of positive ions outside. These excess charges collect in a thin shell tightly against the inner and outer surfaces of the plasma membrane (Figure 6.9), while the bulk of the intracellular and extracellular fluid remains electrically neutral. Unlike the diagrammatic representation, the number of charges separated across the membrane to account for the potential is an infinitesimal fraction of the total charges in both compartments.
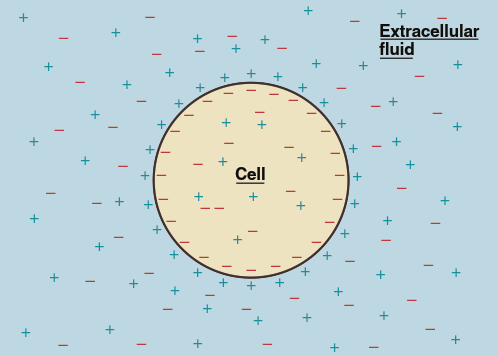
Figure 6.9. The excess positive charges outside the cell and the excess negative charges inside collect in a tight shell against the plasma membrane. In reality, these excess charges are only an extremely small fraction of the total number of ions inside and outside the cell
Table 6.2 lists the concentrations of sodium (Na⁺), potassium (K⁺), and chloride (Cl⁻) ions in the extracellular and intracellular fluid of a representative neuron. Each of these ions shows a 10‑ to 30‑fold concentration difference between the inside and outside of the cell. Although this table might appear to contradict the assertion that the bulk of fluid is electrically neutral, many other ions are present—including Mg²⁺, Ca²⁺, H⁺, HCO₃⁻, HPO₄²⁻, SO₄²⁻, and ionized organic compounds such as amino acids and proteins. When all ions are accounted for, each solution is indeed electrically neutral.
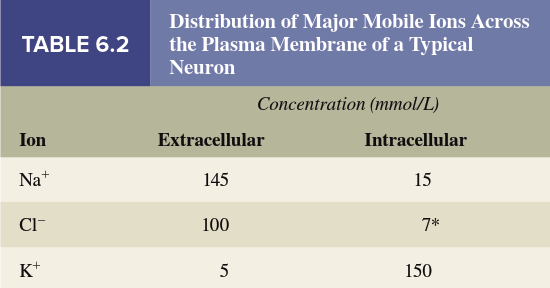
Of the ions that can flow across the membrane and affect its electrical potential, Na⁺, K⁺, and Cl⁻ are present in the highest concentrations, and the membrane permeability to each is independently determined. Na⁺ and K⁺ generally make the most important contributions to generating the resting membrane potential, though in some cells Cl⁻ is also a factor. Notice that Na⁺ and Cl⁻ concentrations are lower inside the cell than outside, while the K⁺ concentration is greater inside. The concentration differences for Na⁺ and K⁺ are established by the sodium‑potassium ATPase pump (Na⁺/K⁺‑ATPase) , which pumps Na⁺ out of the cell and K⁺ into it. The reason for the Cl⁻ distribution varies among cell types, as described later.
The magnitude of the resting membrane potential depends mainly on two factors: (1) differences in specific ion concentrations between intracellular and extracellular fluids; and (2) differences in membrane permeabilities to the different ions, which reflect the number of open channels for each ion in the plasma membrane. A third factor—a direct contribution from ion pumps—has a lesser role. We will examine each of these in detail.
Contribution of Ion Concentration Differences. To understand how concentration differences for Na⁺ and K⁺ create membrane potentials, first consider what happens when the membrane is permeable (has open channels) to only one ion (Figure 6.10). In this hypothetical situation, assume the membrane contains K⁺ channels but no Na⁺ or Cl⁻ channels. Initially, compartment 1 contains 0.15 M NaCl, compartment 2 contains 0.15 M KCl, and no ion movement occurs because the channels are closed (Figure 6.10a). There is no potential difference because the two compartments contain equal numbers of positive and negative ions. However, when these K⁺ channels open, K⁺ diffuses down its concentration gradient from compartment 2 into compartment 1 (Figure 6.10b). After a few potassium ions move, compartment 1 gains an excess of positive charge, leaving an excess of negative charge in compartment 2 (Figure 6.10c), thus creating a potential difference across the membrane.
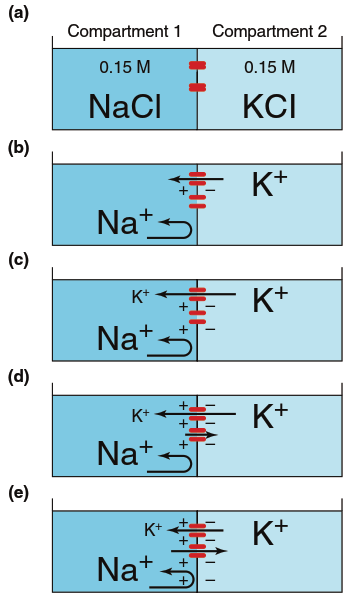
Figure 6.10. Generation of a potential across a membrane due to diffusion of K1 through K1 channels (red). Arrows represent ion movements; as in Figure 4.3, arrow length represents the magnitude of the flux. See the text for a complete explanation of the steps a–e
This introduces another major factor that can cause net ion movement: an electrical potential. As compartment 1 becomes increasingly positive and compartment 2 increasingly negative, the membrane potential begins to influence K⁺ movement. The negative charge of compartment 2 attracts K⁺ back, and the positive charge of compartment 1 repels them (Figure 6.10d). In other words, an electrochemical gradient exists across the membrane for all ions. As long as the flux due to the K⁺ concentration gradient exceeds the flux due to the membrane potential, net K⁺ movement occurs from compartment 2 to compartment 1, and the membrane potential progressively increases. Eventually, the membrane potential becomes negative enough to produce a flux equal but opposite to that produced by the concentration gradient (Figure 6.10e). The membrane potential at which these two fluxes become equal in magnitude but opposite in direction is called the equilibrium potential for that ion (here, K⁺). At the equilibrium potential, there is no net movement of the ion, and the potential undergoes no further change.
The number of ions crossing the membrane to establish this equilibrium potential is insignificant compared to the number originally present, so there is no significant change in K⁺ concentration in either compartment between step (a) and step (e). The magnitude of the equilibrium potential (in mV) for any ion depends on its concentration gradient across the membrane. If concentrations on both sides were equal, the net flux would be zero and the equilibrium potential would also be zero. The larger the concentration gradient, the larger the equilibrium potential, because a greater electrically driven movement of ions is required to balance the movement due to the concentration difference.
Now consider a membrane that contains only Na⁺ channels (Figure 6.11). Sodium ions (Na⁺) will initially move from compartment 1 to compartment 2. When compartment 2 becomes positive relative to compartment 1, the electrical charge difference will begin to drive Na⁺ back, and eventually net movement ceases. Again, at the equilibrium potential, the movement due to the concentration gradient equals but opposes the movement due to the electrical gradient, and an insignificant number of sodium ions actually move. Thus, the equilibrium potential for one ion can differ in magnitude and direction from those for other ions, depending on the concentration gradients between intracellular and extracellular compartments.
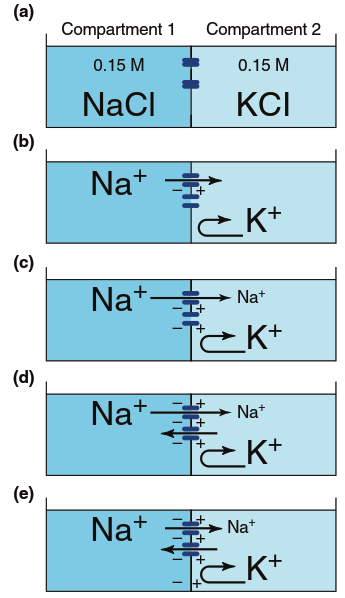
Figure 6.11. Generation of a potential across a membrane due to diffusion of Na+ through Na+ channels (blue). Arrows represent ion movements; as in Figure 4.3, arrow length indicates the magnitude of the flux. So few sodium ions cross the membrane that ion concentrations do not change significantly from step (a) to step (e). See the text for a more complete explanation
Is there a way to predict how much electrical force is required to exactly balance the tendency of an ion to diffuse down its concentration gradient? The Nernst equation describes the equilibrium potential for any ion—that is, the electrical potential necessary to balance a given ionic concentration gradient across a membrane so that the net flux of the ion is zero. The Nernst equation is:

Using the concentration gradients from Table 6.2, the equilibrium potentials for Na⁺ (E_Na) and K⁺ (E_K) are:
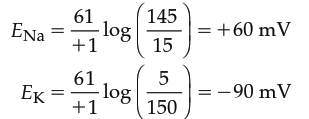
Thus, at these typical concentrations, Na⁺ flux through open channels will tend to bring the membrane potential toward +60 mV, whereas K⁺ flux will bring it toward –90 mV. If the concentration gradients change, the equilibrium potentials will change. The hypothetical situations in Figures 6.10 and 6.11 are useful for understanding how individual permeating ions influence membrane potential, but real cells are far more complicated—many charged molecules contribute to overall electrical properties, and cells are rarely permeable to only a single ion at a time.
Contribution of Different Ion Permeabilities. When channels for more than one type of ion are open in the membrane simultaneously, the permeabilities and concentration gradients for all ions must be considered to account for the membrane potential. For a given concentration gradient, the greater the membrane permeability to one type of ion, the greater that ion’s contribution to the membrane potential. Given the concentration gradients and relative membrane permeabilities (P_ion) for Na⁺, K⁺, and Cl⁻, the resting membrane potential (Vₘ) can be calculated using the Goldman‑Hodgkin‑Katz (GHK) equation:
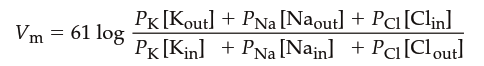
The GHK equation is an expanded version of the Nernst equation that takes into account individual ion permeabilities. Setting the permeabilities of any two ions to zero gives the equilibrium potential for the remaining ion. Note that the Cl⁻ concentrations are reversed compared to Na⁺ and K⁺ (inside concentration in numerator, outside in denominator) because Cl⁻ is an anion and its movement has the opposite effect on the membrane potential. Ion gradients and permeabilities vary widely among different excitable cells, yet the GHK equation can determine the resting membrane potential of any cell if the conditions are known. For example, if the relative permeability values of a cell were P_K = 1, P_Na = 0.04, and P_Cl = 0.45, and the ion concentrations equaled those listed in Table 6.2, the resting membrane potential would be:
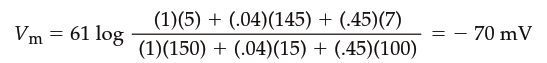
The contributions of Na⁺, K⁺, and Cl⁻ to the overall membrane potential are thus a function of their concentration gradients and relative permeabilities. The concentration gradients determine their equilibrium potentials, and the relative permeability determines how strongly the resting membrane potential is influenced toward those potentials. In mammalian neurons, K⁺ permeability may be as much as 100 times greater than that for Na⁺ and Cl⁻, so neuronal resting membrane potentials are typically fairly close to the equilibrium potential for K⁺ (Figure 6.12). The value of the Cl⁻ equilibrium potential is also near the resting membrane potential in many neurons, but Cl⁻ has minimal importance in determining neuronal resting membrane potentials compared to K⁺ and Na⁺.
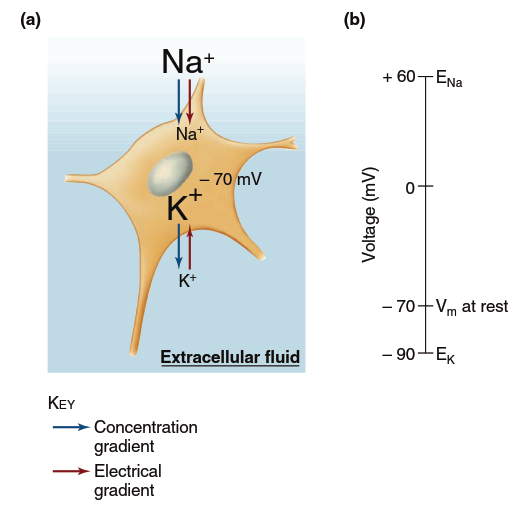
Figure 6.12. Forces influencing sodium and potassium ions at the resting membrane potential (Vm). (a) At a resting membrane potential of 270 mV, both the concentration and electrical gradients favor inward movement of Na+, whereas the K+ concentration and electrical gradients are in opposite directions. (b) The greater permeability and movement of K+ maintain the resting membrane potential at a value near EK.
In summary, the resting potential is generated largely by the movement of K⁺ out of the cell down its concentration gradient through constitutively open K⁺ channels (called leak channels or ungated channels to distinguish them from gated channels). This makes the inside of the cell negative relative to the outside. Even though K⁺ flux has more impact on the resting membrane potential than Na⁺ flux, the resting membrane potential is not equal to the K⁺ equilibrium potential because a small number of open leak channels for Na⁺ pulls the membrane potential slightly toward the Na⁺ equilibrium potential. Thus, at rest, ion channels allow net movement of both Na⁺ into the cell and K⁺ out of the cell.
Contribution of Ion Pumps. Over time, the concentrations of intracellular sodium and potassium ions do not change because of the action of the Na⁺/K⁺‑ATPase pump. In a resting cell, the number of ions the pump moves equals the number that leak down their electrochemical gradients. As long as the concentration gradients remain stable and the ion permeabilities of the plasma membrane do not change, the electrical potential across the resting membrane remains constant. Thus far, we have described the membrane potential as due purely to the passive movement of ions down their electrochemical gradients, with the concentration gradients maintained by membrane pumps. However, the Na⁺/K⁺‑ATPase pump not only maintains these concentration gradients but also establishes them in the first place.
Additionally, the pump helps establish the membrane potential more directly. The Na⁺/K⁺‑ATPase moves three Na⁺ out of the cell for every two K⁺ it brings in. This unequal transport of positive ions makes the inside of the cell more negative than it would be from ion diffusion alone. When a pump moves net charge across the membrane and contributes directly to the membrane potential, it is known as an electrogenic pump. In most cells, the electrogenic contribution to the membrane potential is quite small. Nevertheless, the pump always makes an essential indirect contribution because it maintains the concentration gradients that result in ion diffusion and charge separation.
Summary of the Development of a Resting Membrane Potential. Figure 6.13 summarizes the development of a resting membrane potential in three conceptual steps. First, the action of the Na⁺/K⁺‑ATPase pump sets up the concentration gradients for Na⁺ and K⁺ (Figure 6.13a). These concentration gradients determine the equilibrium potentials for the two ions—that is, the value to which each ion would bring the membrane potential if it were the only permeating ion. Simultaneously, the pump has a small electrogenic effect due to pumping three Na⁺ out for every two K⁺ in. Next, initially there is a greater flux of K⁺ out of the cell than Na⁺ into the cell (Figure 6.13b) because in a resting membrane the permeability (more leak channels) is greater to K⁺ than to Na⁺. Because there is greater net efflux than influx of positive ions during this step, a significant negative membrane potential develops, approaching the K⁺ equilibrium potential. In the steady‑state resting neuron, the flux of ions across the membrane reaches a dynamic balance (Figure 6.13c). Because the membrane potential is not equal to the equilibrium potential for either ion, there is a small but steady leak of Na⁺ into the cell and K⁺ out of the cell. The concentration gradients do not dissipate over time, however, because ion movement by the Na⁺/K⁺‑ATPase pump exactly balances the rate at which the ions leak in the opposite direction.
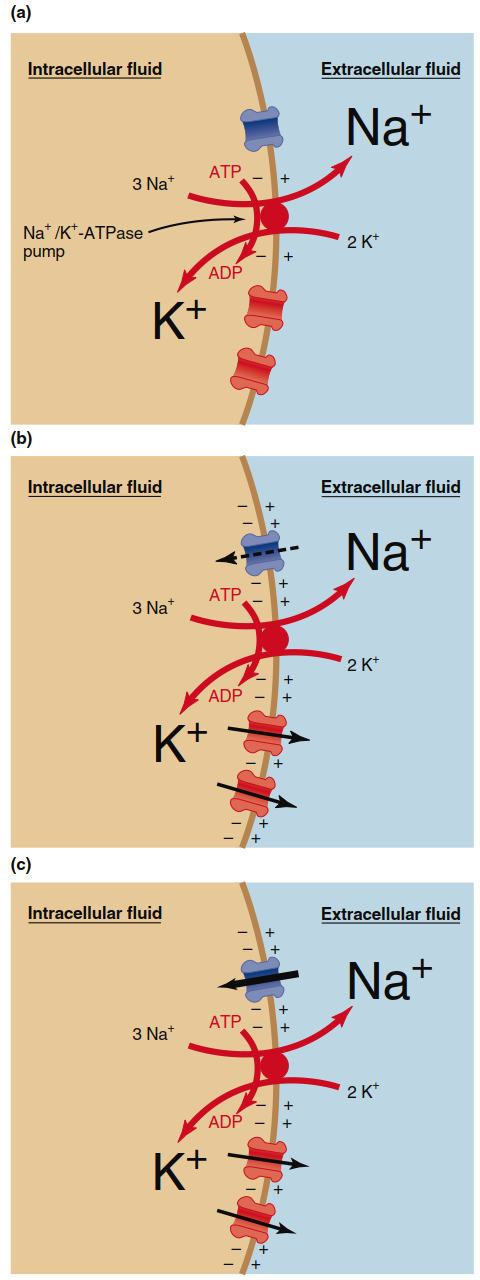
Figure 6.13. Summary of steps establishing the resting membrane potential. (a) An Na+/K+-ATPase pump establishes concentration gradients and generates a small negative potential. (b) Greater net movement of K+ than Na+ makes the membrane potential more negative on the inside. (c) At a steady negative resting membrane potential, ion fluxes through the channels and pump balance each other.
Chloride Ions and Resting Potential. Now let us return to the behavior of chloride ions in excitable cells. The plasma membranes of many cells also have Cl⁻ channels but do not contain chloride ion pumps. Therefore, in these cells, Cl⁻ concentrations simply shift until the equilibrium potential for Cl⁻ equals the resting membrane potential. In other words, the negative membrane potential determined by Na⁺ and K⁺ moves Cl⁻ out of the cell, making the intracellular Cl⁻ concentration lower than that outside. This concentration gradient produces a diffusion of Cl⁻ back into the cell that exactly opposes the movement out due to the electrical potential. In contrast, some cells have a nonelectrogenic active‑transport system that moves Cl⁻ out of the cell, generating a strong concentration gradient. In these cells, the Cl⁻ equilibrium potential is negative to the resting membrane potential, and net Cl⁻ diffusion into the cell contributes to the excess negative charge inside the cell; that is, net Cl⁻ diffusion makes the membrane potential more negative than it would be if only Na⁺ and K⁺ were involved.
Date added: 2026-07-14; views: 4;