Cell Signaling and Receptor Types: A Scientific Guide to Signal Transduction
In Chapter 1, you learned that several classes of chemical messengers can communicate a signal from one cell to another. These messengers include molecules such as neurotransmitters and paracrine substances, whose signals are mediated rapidly and over short distances. Other messengers, such as hormones, communicate over greater distances and, in some cases, more slowly. Regardless of the chemical messenger type, the receiving cell must possess a mechanism to detect the signal's presence. Once a cell detects a signal, a process is required to transduce that signal into a physiologically meaningful response, such as cell division following growth-promoting signals.
The first step in any intercellular chemical messenger action is the binding of the messenger to specific target-cell proteins known as receptors (or receptor proteins). In general biochemical terms, a chemical messenger functions as a ligand, and the receptor contains a binding site specific for that ligand. The binding of a messenger to its receptor induces a conformational change (tertiary structure alteration; see Figure 2.17) in the receptor, thereby activating it. This activation initiates a sequence of cellular events leading to the cell's response, a process termed signal transduction. The "signal" refers to receptor activation, while "transduction" describes the transformation of a stimulus into a cellular response. This section examines general features common to many receptors, describes receptor-ligand interactions, and provides examples of receptor regulation.
Types of Receptors. Receptors that bind intercellular chemical messengers are proteins or glycoproteins located either in the cell's plasma membrane or inside the cell (in the cytosol or nucleus). The plasma membrane represents the more common location because numerous messengers are water-soluble and cannot diffuse across the lipid-rich (hydrophobic) plasma membrane. In contrast, a smaller number of lipid-soluble messengers can diffuse through membranes to bind their intracellular receptors.
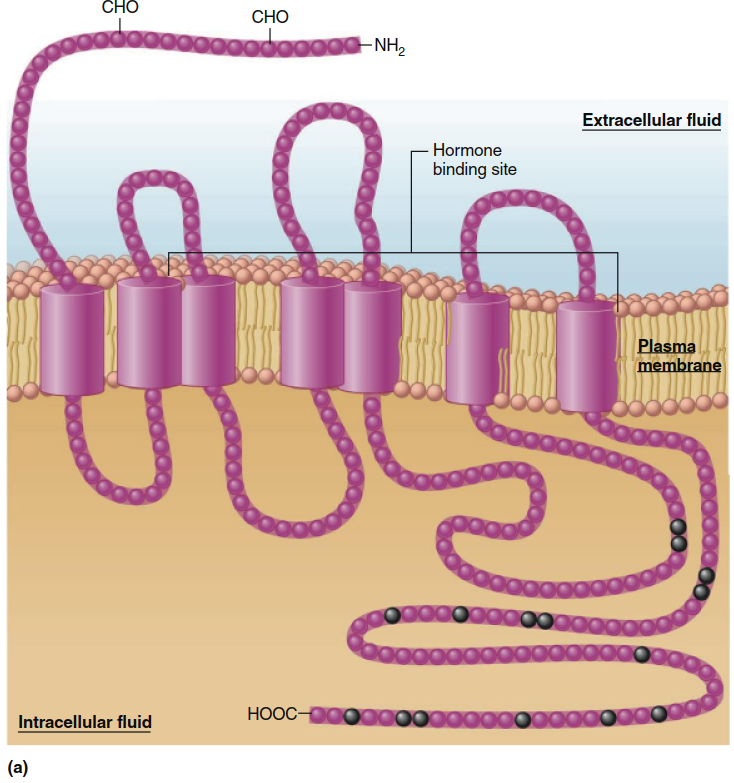
Plasma Membrane Receptors (illustrated in Figure 5.1a) are transmembrane proteins that span the entire membrane thickness. Like other transmembrane proteins, a plasma membrane receptor possesses hydrophobic segments within the membrane, one or more hydrophilic segments extending into the extracellular fluid, and additional hydrophilic segments projecting into the intracellular fluid. Arriving chemical messengers bind to the extracellular regions of the receptor, while the intracellular regions participate in signal transduction events.
Intracellular Receptors (shown in Figure 5.1b) are not located in membranes but exist in either the cytosol or the cell nucleus and have a distinctly different structure. Like plasma membrane receptors, however, they possess a segment that binds the messenger and other segments that function as regulatory sites. Additionally, intracellular receptors contain a DNA-binding segment, which distinguishes them from plasma membrane receptors. This represents a key distinction: plasma membrane receptors can transduce signals without interacting with DNA, whereas all intracellular receptors transduce signals through interactions with genes.
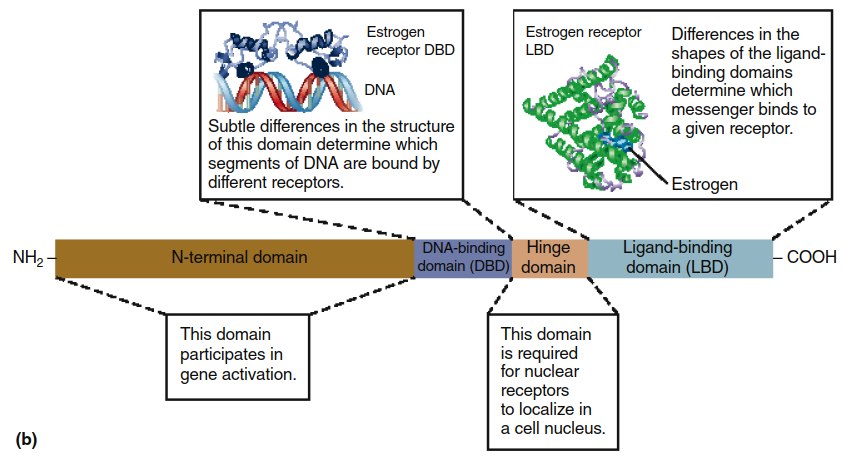
Figure 5.1. The two major classes of receptors for chemical messengers. (a) Structure of a typical transmembrane receptor. The seven clusters of amino acids embedded in the phospholipid bilayer represent hydrophobic portions of the protein’s alpha helix (shown here as cylinders). Note that the binding site for the hormone includes several of the segments that extend into the extracellular fluid. Portions of the extracellular segments can be linked to carbohydrates (CHO). The amino acids denoted by black circles represent some of the sites at which intracellular enzymes can phosphorylate, and thereby regulate, the receptor. (b) Schematic representation of the structural features of a typical nuclear receptor. The actual structures for segments of these receptors are known and are shown here for the human estrogen (a steroid hormone) receptor. (Note: The segments of nuclear receptors that perform different functions are known as “domains.”)
Interactions Between Receptors and Ligands. Four major features define the interactions between receptors and their ligands: specificity, affinity, saturation, and competition (see Table 5.1 for a glossary of receptor terminology).

Specificity refers to the binding of a chemical messenger to its receptor, which initiates events leading to the cellular response. The existence of receptors explains a crucial characteristic of intercellular communication—specificity. Although a given chemical messenger may contact many different cell types, it influences only certain cell types because cells differ in the receptor types they possess. Only specific cell types—sometimes just one—express the particular receptor required to bind a given chemical messenger (Figure 5.2).
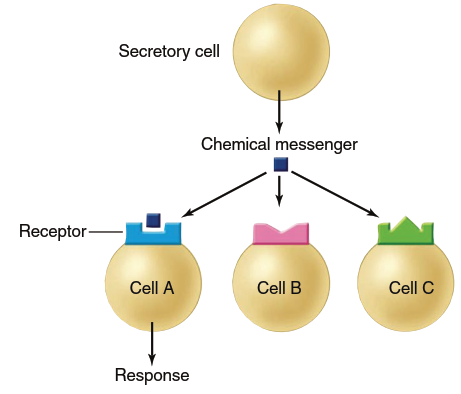
Figure 5.2. Specificity of receptors for chemical messengers. Only cell A has the appropriate receptor for this chemical messenger; therefore, it is the only one among the group that is a target cell for the messenger
Even when different cell types possess receptors for the same messenger, their responses to that messenger may differ substantially. For example, the neurotransmitter norepinephrine causes smooth muscle contraction in certain blood vessels but, via the same receptor type, inhibits insulin secretion from the pancreas. Thus, the receptor functions as a molecular switch that elicits the cell's response when activated by messenger binding. Just as identical switches can operate a light or a radio, a single receptor type can produce different responses in different cell types.
Affinity determines the degree to which a particular messenger binds to its receptor (summarized in Figure 5.3). A receptor with high affinity will bind at lower messenger concentrations than a receptor with low affinity (refer back to Figure 3.36). Differences in receptor affinity for their ligands have important implications for therapeutic drugs in treating illness; receptors with high affinity for a ligand require much less ligand (a lower dose) to become activated.
Saturation of receptors (described in Chapter 3 for ligand-protein binding) applies fully here (see Figure 5.3). A cell's response to a messenger increases as the extracellular messenger concentration increases because the number of occupied receptors grows. However, there is an upper limit to this responsiveness because only a finite number of receptors are available, and they become fully saturated at some point.
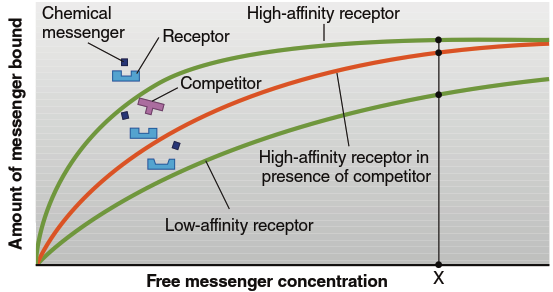
Figure 5.3. Characteristics of receptors binding to messengers. The receptors with high affinity will have more bound messenger at a given messenger concentration (e.g., concentration X). The presence of a competitor will decrease the amount of messenger bound, until at very high concentrations the receptors become saturated with messenger. Note in the illustration that the low-affinity receptor in this case has a slightly different shape in its ligand-binding region compared to the high-affinity receptor. Also note the similarity in parts of the shapes of the natural messenger and its competitor
Competition refers to the ability of a molecule to compete with a natural ligand for binding to its receptor. Competition typically occurs with messengers having structural similarities and underlies the action of many drugs (see Figure 5.3). When researchers or physicians wish to interfere with a particular messenger's action, they can administer competing molecules structurally similar enough to bind to that messenger's receptors. However, these competing molecules differ sufficiently from the native ligand that they cannot activate the receptor, thereby blocking the endogenous messenger from binding without inducing signal transduction.
The general term for a compound that blocks a chemical messenger's action is antagonist; when an antagonist works by competing for the binding site, it is known as a competitive antagonist. One example is beta-adrenergic receptor blockers (also called beta-blockers), used in treating high blood pressure and other diseases. Beta-blockers compete with epinephrine and norepinephrine for binding to beta-adrenergic receptors. Because epinephrine and norepinephrine normally increase blood pressure (Chapter 12), beta-blockers decrease blood pressure by acting as competitive antagonists. Antihistamines are another example, useful in treating allergic symptoms from excess histamine secretion from mast cells (Chapter 18). Antihistamines are competitive antagonists that block histamine from binding to its receptors on mast cells, preventing allergic responses.
Some drugs that compete with natural ligands for a particular receptor type do activate the receptor and trigger the cellular response exactly as if the endogenous chemical messenger had bound. Such drugs, known as agonists, are used therapeutically to mimic the messenger's action. For example, the common decongestants phenylephrine and oxymetazoline (found in many nasal sprays) mimic epinephrine's action on alpha-adrenergic receptors in blood vessels. When alpha-adrenergic receptors are activated, the smooth muscles of inflamed, dilated nasal blood vessels contract, resulting in vasoconstriction that helps open nasal passages and decrease fluid leakage.
Regulation of Receptors. Receptors are themselves subject to physiological regulation. The number of receptors a cell possesses, or the affinity of receptors for their specific messenger, can be increased or decreased in certain systems. An important example is down-regulation: when a high extracellular messenger concentration is maintained over time, the total number of target cell receptors for that messenger may decrease. Down-regulation reduces target cell responsiveness to frequent or intense stimulation—desensitizing them—and represents a local negative feedback mechanism.
Down-regulation occurs because there is continuous synthesis and degradation of receptors. The primary mechanism of down-regulation for plasma membrane receptors is internalization. Messenger binding to its receptor can stimulate internalization of the messenger-receptor complex, which is taken into the cell via receptor-mediated endocytosis. This process increases the rate of receptor degradation inside the cell. Consequently, at elevated messenger concentrations, the number of plasma membrane receptors gradually decreases during down-regulation.
The opposite change, called up-regulation, also occurs. Cells exposed for prolonged periods to very low messenger concentrations may develop many more receptors for that messenger, thereby developing increased sensitivity to it. The greater the number of receptors available to bind a ligand, the greater the likelihood that binding will occur. For example, when nerves to a muscle are damaged, neurotransmitter delivery decreases or ceases entirely. Over time, under these conditions, the muscle will contract in response to much smaller amounts of neurotransmitter than normal because the receptors have been up-regulated, resulting in increased sensitivity.
One mechanism of up-regulation involves recruitment to the plasma membrane of intracellular vesicles containing numerous receptor proteins within their membranes. These vesicles fuse with the plasma membrane, thereby inserting their receptors into the membrane. Receptor regulation in both directions (up-regulation and down-regulation) is an excellent example of the general physiological principle of homeostasis, as it acts to return signal strength toward normal when messenger concentrations vary above or below normal levels.
Date added: 2026-07-14; views: 6;