Cyclic AMP, Calcium, and Eicosanoids: Key Signal Transduction Pathways in Cell Communication
Cyclic AMP Pathway and Its Amplification Cascade. In the cyclic AMP (cAMP) pathway (Figure 5.6), activation of the receptor by binding of the first messenger (for example, the hormone epinephrine) allows the receptor to activate its associated G protein, known as Gs (the subscript s denotes “stimulatory”). This activation causes Gs to stimulate its effector protein, the plasma membrane enzyme called adenylyl cyclase (also known as adenylate cyclase). The activated adenylyl cyclase, with its catalytic site on the cytosolic surface of the plasma membrane, catalyzes the conversion of cytosolic ATP to cyclic 3',5'-adenosine monophosphate (cAMP) (Figure 5.7). Cyclic AMP then acts as a second messenger (see Figure 5.6), diffusing throughout the cell to trigger the sequence of events leading to the cell’s ultimate response to the first messenger.
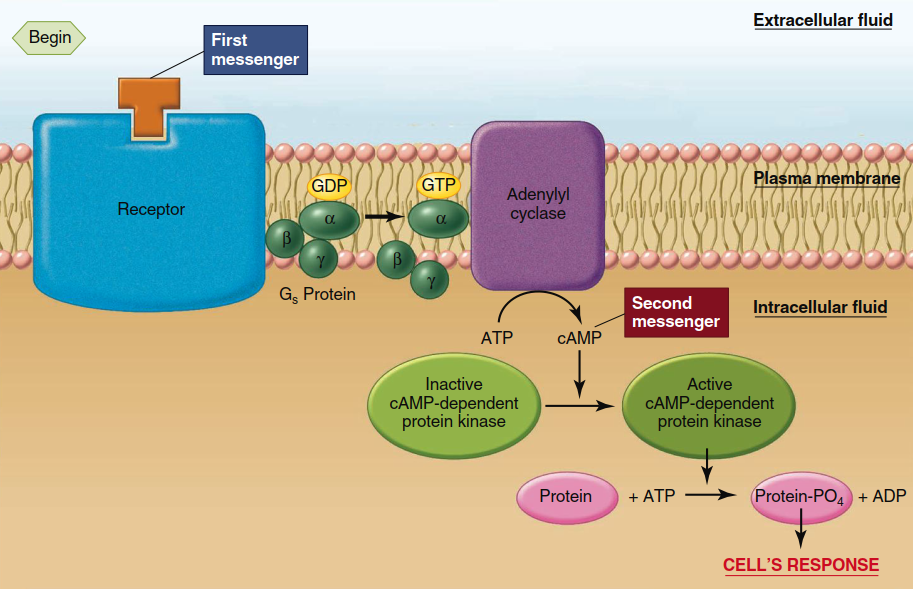
Figure 5.6. Cyclic AMP second-messenger system. Not shown in the figure is the existence of another regulatory protein, Gi, which certain receptors can react with to cause inhibition of adenylyl cyclase
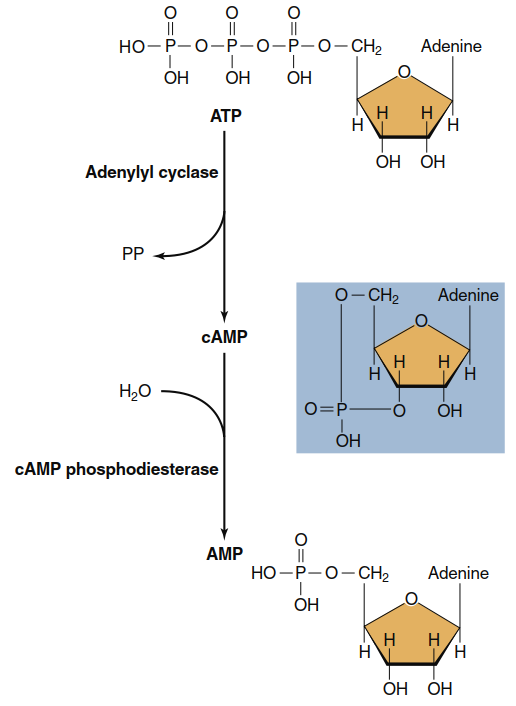
Figure 5.7. Formation and breakdown of cAMP. ATP is converted to cAMP by the action of the plasma membrane enzyme adenylyl cyclase. cAMP is inactivated by the cytosolic enzyme cAMP phosphodiesterase, which converts cAMP into the noncyclized form AMP
The action of cAMP eventually terminates when it is broken down to AMP, a reaction catalyzed by the enzyme cAMP phosphodiesterase (see Figure 5.7). This enzyme is also subject to physiological control, so the cellular concentration of cAMP can be changed either by altering the rate of its messenger-mediated synthesis or the rate of its phosphodiesterase-mediated breakdown. Caffeine and theophylline (the active ingredients of coffee and tea) are widely consumed stimulants that work partly by inhibiting cAMP phosphodiesterase activity, thereby prolonging the actions of cAMP within cells. In many cells, such as those of the heart, an increased concentration of cAMP triggers an increase in function (for example, an increase in heart rate).
What does cAMP actually do inside the cell? It binds to and activates an enzyme known as cAMP-dependent protein kinase, also called protein kinase A (see Figure 5.6). Recall that protein kinases phosphorylate other proteins—often enzymes—by transferring a phosphate group to them. The changes in the activity of proteins phosphorylated by cAMP-dependent protein kinase bring about a cell’s response (secretion, contraction, and so on). Again, each of the various protein kinases that participate in the multiple signal transduction pathways has its own specific substrates.
In essence, the activation of adenylyl cyclase by the Gs protein initiates an “amplification cascade” of events that converts proteins in sequence from inactive to active forms. Figure 5.8 illustrates the benefit of such a cascade. While active, a single enzyme molecule can transform not one but many substrate molecules—let us say 100—into product. Therefore, one active molecule of adenylyl cyclase may catalyze the generation of 100 cAMP molecules (and thus 100 activated cAMP-dependent protein kinase A molecules). At each of the two subsequent enzyme-activation steps, another 100-fold amplification occurs, so a single molecule of the first messenger could cause the generation of 1 million product molecules. This helps explain how hormones and other messengers can be effective at extremely low extracellular concentrations. For example, one molecule of epinephrine can cause the liver to generate and release 10⁸ molecules of glucose.
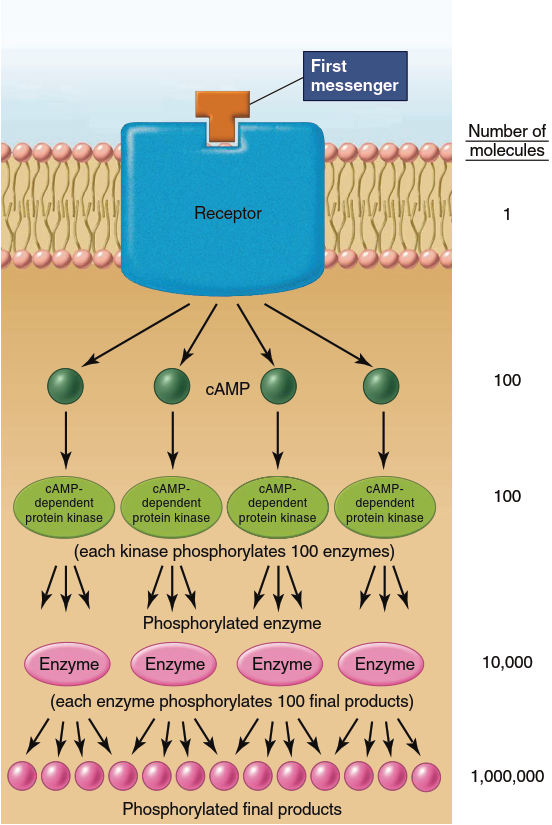
Figure 5.8. Example of signal amplification. In this example, a single molecule of a first messenger results in 1 million final products. Other second-messenger pathways have similar amplification processes. The steps between receptor activation and cAMP generation are omitted for simplicity
In addition, activated cAMP-dependent protein kinase can diffuse into the cell nucleus, where it can phosphorylate a protein that then binds to specific regulatory regions of certain genes (such genes are said to be cAMP-responsive). Therefore, the effects of cAMP can be rapid and independent of changes in gene activity (as in epinephrine and glucose production) or slower and dependent upon the formation of new gene products. How can cAMP’s activation of a single molecule (cAMP-dependent protein kinase) be common to such a variety of biochemical sequences and cell responses? The answer is that cAMP-dependent protein kinase can phosphorylate a large number of different proteins (Figure 5.9), thereby exerting multiple actions within a single cell and different actions in different cells.
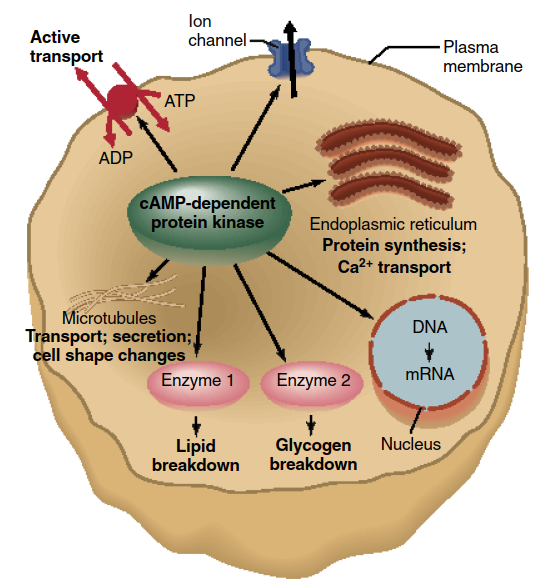
Figure 5.9. The variety of cellular responses induced by cAMP is due mainly to the fact that activated cAMP-dependent protein kinase can phosphorylate many different proteins, activating or inhibiting them. In this figure, the protein kinase is shown phosphorylating seven different proteins—a microtubular protein, an ATPase, an ion channel, a protein in the endoplasmic reticulum, a protein involved in stimulating the transcription of a gene into mRNA, and two enzymes
For example, epinephrine acts via the cAMP pathway on adipose cells to stimulate the breakdown of triglyceride, a process mediated by a particular phosphorylated enzyme chiefly expressed in adipose cells. In the liver, epinephrine acts via cAMP to stimulate both glycogenolysis and gluconeogenesis, processes mediated by phosphorylated enzymes that differ from those expressed in adipose cells. Whereas phosphorylation mediated by cAMP-dependent protein kinase activates certain enzymes, it inhibits others. For instance, the enzyme catalyzing the rate-limiting step in glycogen synthesis is inhibited by phosphorylation, explaining how epinephrine inhibits glycogen synthesis while simultaneously stimulating glycogen breakdown.
Not mentioned thus far is that receptors for some first messengers, upon activation, inhibit adenylyl cyclase, resulting in less cAMP generation. This occurs because these receptors are associated with a different G protein known as Gi (the subscript i denotes “inhibitory”). Activation of Gi causes inhibition of adenylyl cyclase, decreasing the cellular cAMP concentration and thereby reducing phosphorylation of key proteins. Many cells express both stimulatory (Gs) and inhibitory (Gi) G proteins in their membranes, providing a means of tightly regulating intracellular cAMP concentrations. This common feature highlights the principle that most physiological functions are controlled by multiple regulatory systems, often working in opposition, allowing fine-tuning of cellular responses. Finally, as indicated in Figure 5.9, cAMP-dependent protein kinase can phosphorylate certain plasma membrane ion channels, causing them to open or close. This indirect G-protein gating of ion channels utilizes a second-messenger pathway, distinct from the direct action of a G protein on an ion channel. Table 5.2 summarizes the three ways by which receptor activation leads to opening or closing of ion channels, causing changes in membrane potential.

Phospholipase C, Diacylglycerol, and Inositol Trisphosphate Pathway. In this system, a G protein called Gq is activated by a receptor bound to a first messenger. Activated Gq then activates a plasma membrane effector enzyme called phospholipase C. This enzyme catalyzes the breakdown of a plasma membrane phospholipid known as phosphatidylinositol bisphosphate (PIP₂) to diacylglycerol (DAG) and inositol trisphosphate (IP₃) (Figure 5.10). Both DAG and IP₃ then function as second messengers but in very different ways.
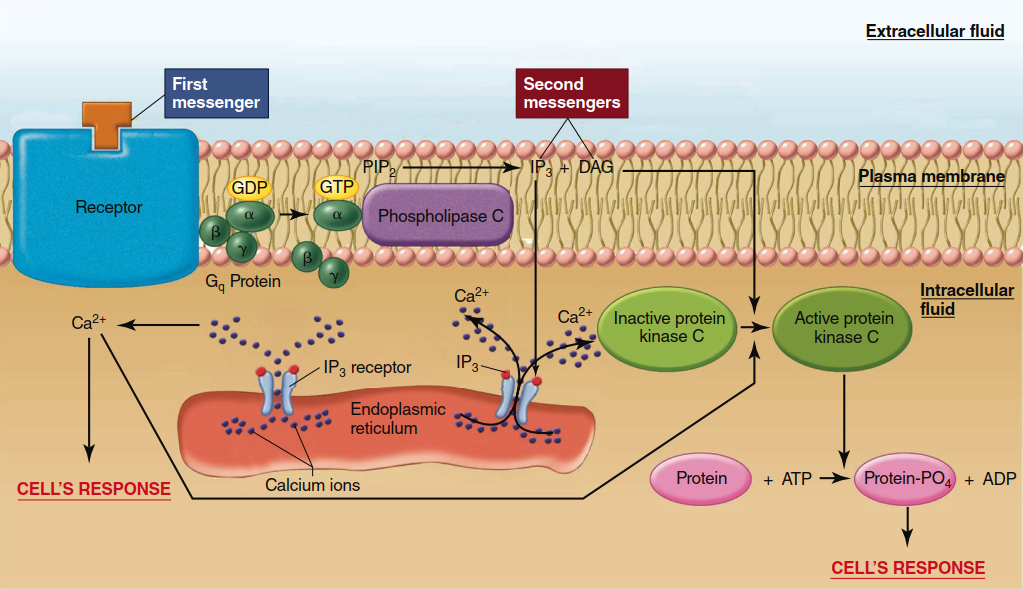
Figure 5.10. Mechanism by which an activated receptor stimulates the enzymatically mediated breakdown of PIP2 to yield IP3 and DAG. IP3 then binds to a receptor on the endoplasmic receptor. This receptor is a ligand-gated ion channel that, when opened, allows the release of Ca2+ from the endoplasmic reticulum into the cytosol. Together with DAG, Ca2+ activates protein kinase C.
DAG activates members of a family of related protein kinases known collectively as protein kinase C, which, similarly to cAMP-dependent protein kinase, then phosphorylates a large number of other proteins, leading to the cell’s response. IP₃, in contrast, does not exert its second-messenger function by directly activating a protein kinase. Rather, cytosolic IP₃ binds to receptors located on the endoplasmic reticulum; these receptors are ligand-gated Ca²⁺ channels that open when bound to IP₃. Because the concentration of Ca²⁺ is much greater in the endoplasmic reticulum than in the cytosol, Ca²⁺ diffuses out of this organelle into the cytosol, significantly increasing the cytosolic Ca²⁺ concentration. This increased Ca²⁺ concentration then continues the sequence of events leading to the cell’s response to the first messenger. Notably, one action of Ca²⁺ is to help activate some forms of protein kinase C (hence the name “C” for calcium).
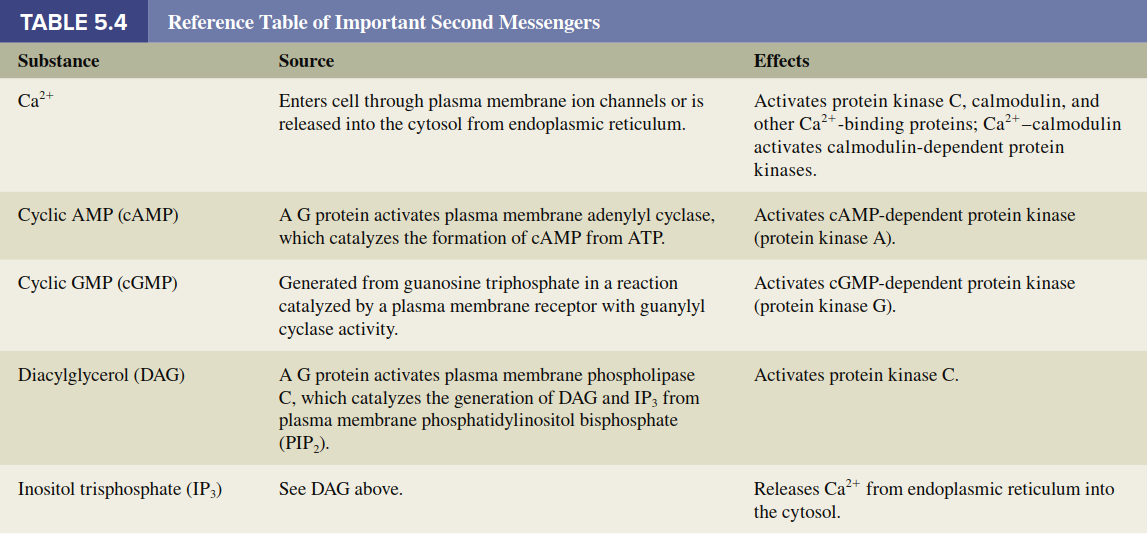
Calcium Ion as a Second Messenger. The calcium ion (Ca²⁺) functions as a second messenger in a great variety of cellular responses to both chemical and electrical stimuli. The physiology of Ca²⁺ as a second messenger requires analysis of two broad questions: (1) How do stimuli cause the cytosolic Ca²⁺ concentration to increase? (2) How does the increased Ca²⁺ concentration elicit the cells’ responses? By means of active-transport systems in the plasma membrane and membranes of certain cell organelles, Ca²⁺ is maintained at an extremely low concentration in the cytosol. Consequently, there is always a large electrochemical gradient favoring diffusion of Ca²⁺ into the cytosol via Ca²⁺ channels found in both the plasma membrane and the endoplasmic reticulum. A stimulus can alter this steady state by influencing the active-transport systems and/or ion channels, resulting in a change in cytosolic Ca²⁺ concentration. The most common ways that receptor activation increases cytosolic Ca²⁺ are summarized in the top part of Table 5.3.

Now we turn to how the increased cytosolic Ca²⁺ concentration elicits cellular responses (see bottom of Table 5.3). The common denominator of Ca²⁺ actions is its ability to bind to various cytosolic proteins, altering their conformation and thereby activating their function. One of the most important of these is a protein found in all cells known as calmodulin (Figure 5.11). On binding with Ca²⁺, calmodulin changes shape, allowing Ca²⁺-calmodulin to activate or inhibit a large variety of enzymes and other proteins, many of them protein kinases. Activation or inhibition of these calmodulin-dependent protein kinases leads, via phosphorylation, to activation or inhibition of proteins involved in the cell’s ultimate responses to the first messenger. However, calmodulin is not the only intracellular protein influenced by Ca²⁺ binding. For example, in certain types of muscle, Ca²⁺ binds to a protein called troponin to initiate contraction. For reference, Table 5.4 summarizes the production and functions of the major second messengers described in this chapter.
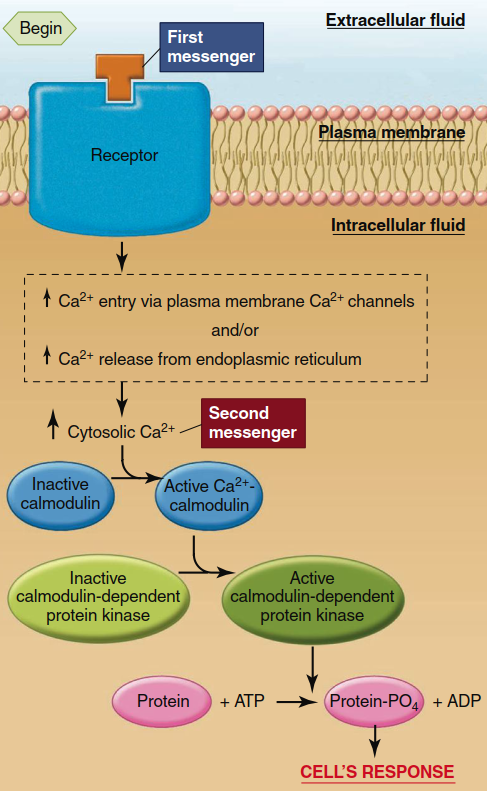
Figure 5.11. Ca2+, calmodulin, and the calmodulin-dependent protein kinase system. (There are multiple calmodulin-dependent protein kinases.) Table 5.3 summarizes the mechanisms for increasing cytosolic Ca2+ concentration
Other Messengers: Eicosanoids. In a few places in this text, you will learn about messengers that are not as readily classified as those described above. Among these are the eicosanoids. The eicosanoids are a family of molecules produced from the polyunsaturated fatty acid arachidonic acid, which is present in plasma membrane phospholipids. The eicosanoids include the cyclic endoperoxides, the prostaglandins, the thromboxanes, and the leukotrienes (Figure 5.12). They are generated in many kinds of cells in response to different extracellular signals, including growth factors, immune defense molecules, and even other eicosanoids. Thus, eicosanoids may act as both extracellular and intracellular messengers, depending on the cell type.
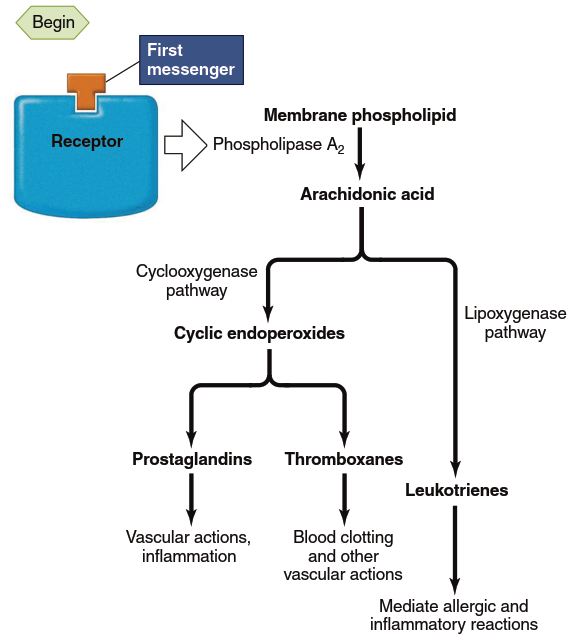
Figure 5.12. Pathways for eicosanoid synthesis and some of their major functions. Phospholipase A2 is the one enzyme common to the formation of all the eicosanoids; it is the site at which stimuli act. Anti-inflammatory steroids inhibit phospholipase A2. The step mediated by cyclooxygenase is inhibited by aspirin and other nonsteroidal anti-inflammatory drugs (NSAIDs). There are also drugs available that inhibit the lipoxygenase enzyme, thereby blocking the formation of leukotrienes. These drugs may be helpful in controlling asthma, in which excess leukotrienes have been implicated in the allergic and inflammatory components of the disease
The synthesis of eicosanoids begins when an appropriate stimulus (hormone, neurotransmitter, paracrine substance, drug, or toxic agent) binds its receptor and activates phospholipase A₂, an enzyme localized to the plasma membrane of the stimulated cell. As shown in Figure 5.12, this enzyme splits off arachidonic acid from the membrane phospholipids. The arachidonic acid can then be metabolized by two pathways: one initiated by the enzyme cyclooxygenase (COX) leading to cyclic endoperoxides, prostaglandins, and thromboxanes; the other initiated by the enzyme lipoxygenase leading to leukotrienes. Within both pathways, synthesis of specific eicosanoids is enzyme-mediated, so the eicosanoid-pathway enzymes expressed in a particular cell determine which eicosanoids the cell synthesizes.
Each major eicosanoid subdivision contains more than one member. On the basis of structural differences, molecules within each subdivision are designated by a letter—for example, PGA and PGE for prostaglandins of the A and E types—which may then be further subdivided (e.g., PGE₂). Once synthesized, eicosanoids may in some cases act as intracellular messengers, but more often they are released immediately and act locally. For this reason, eicosanoids are usually categorized as paracrine and autocrine substances. After they act, they are quickly metabolized by local enzymes to inactive forms. Eicosanoids exert a wide array of effects, particularly on blood vessels and in inflammation, many of which will be described in future chapters.
Certain drugs influence the eicosanoid pathway and are among the most commonly used worldwide. Aspirin, for example, inhibits cyclooxygenase and therefore blocks the synthesis of endoperoxides, prostaglandins, and thromboxanes. Aspirin and other drugs that block cyclooxygenase are collectively termed nonsteroidal anti-inflammatory drugs (NSAIDs). Their major uses are to reduce pain, fever, and inflammation. The term “nonsteroidal” distinguishes them from synthetic glucocorticoids (analogs of steroid hormones made by the adrenal glands) used in large doses as anti-inflammatory drugs; these steroids inhibit phospholipase A₂ and therefore block the production of all eicosanoids.
Cessation of Activity in Signal Transduction Pathways. Once initiated, signal transduction pathways are eventually shut off to prevent chronic overstimulation of a cell, which can be detrimental. The key event is usually the cessation of receptor activation. Responses to messengers are transient events that persist only briefly and subside when the receptor is no longer bound to the first messenger. A major way that receptor activation ceases is by a decrease in the concentration of first-messenger molecules in the region of the receptor, which occurs as enzymes metabolize the first messenger, as the first messenger is taken up by adjacent cells, or as it simply diffuses away.
In addition, receptors can be inactivated in at least three other ways: (1) The receptor becomes chemically altered (usually by phosphorylation), which may decrease its affinity for a first messenger, causing the messenger to be released; (2) phosphorylation of the receptor may prevent further G-protein binding to the receptor; and (3) plasma membrane receptors may be removed when the combination of first messenger and receptor is taken into the cell by endocytosis. These processes are physiologically controlled. For example, in many cases the inhibitory phosphorylation of a receptor is mediated by a protein kinase that was initially activated in response to the first messenger, constituting negative feedback.
This concludes our description of the basic principles of signal transduction pathways. It is essential to recognize that these pathways do not exist in isolation but may be active simultaneously in a single cell, undergoing complex interactions. This is possible because a single first messenger may trigger changes in more than one pathway, and importantly, because many different first messengers may simultaneously influence a cell. Moreover, a great deal of “cross talk” can occur at one or more levels among the various signal transduction pathways. For example, active molecules generated in the cAMP pathway can alter the activity of receptors and signaling molecules generated by other pathways.
Date added: 2026-07-14; views: 3;