Intracellular membranes. Illustration
The intracellular membranes are a varied series of separators with very different structures, compositions and functions. The importance of the membranes inside a cell is once again to create protected and restricted environments where the various cellular components are more likely to interact with each other or where large quantities of solutes in the form of chemical potential energy can be captured (Figure 2.14). Two classic examples are the membranous systems that make up the nuclear envelope and the mitochondria.
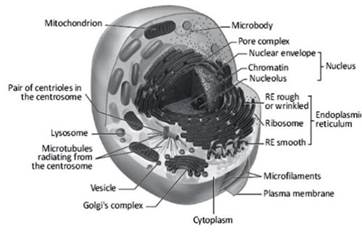
Figure 2.14. The interior of a generic cell and the intracellular membrane complex
The nuclear envelope consists of a double membrane surrounding the genetic material and in eukaryotic cells, it has essentially a containing and protective function (Figure 2.15). There is free communication between nucleoplasm and cytoplasm for ions and small solutes.
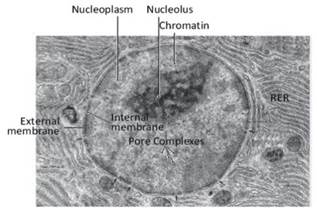
Figure 2.15. Electron micrograph of a portion of a cell. The cell nucleus, the external and internal membranes forming the nuclear envelope, some pore complexes on the nuclear envelope, the section of some pore complexes (black arrows), the wrinkled endoplasmic reticulum (RER) and some cytoplasmic organelles, probably mitochondria, are visible
The genesis of the nuclear envelope, which is a specialization of the endoplasmic reticulum, is unclear; in fact, the outer membrane has ribosomes on its surface and evident continuity with the reticulum cisternae (Figure 2.16). On the contrary, the inner membrane is in close contact with a dense protein network composed of type A and type B laminin, which acts as an anchor for the DNA when it is in its interphasic form.
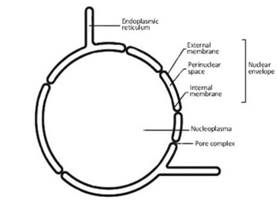
Figure 2.16. Structural composition of the nuclear envelope
Various functions are ascribed to the nuclear envelope. There is experimental evidence that perinuclear cisternae, i.e., the nuclear envelope and the associated wrinkled endoplasmic latticework, are able to accumulate calcium ions and control their concentration in the nucleoplasm.
This task is very important because fluctuations in the concentration of calcium ions can be very harmful to the genetic material: a prolonged increase in the concentration of this ion causes the activation of enzymes such as endonucleases which, by fragmenting DNA, are involved in the apoptotic process.
A very peculiar structure of the nuclear envelope and object of countless studies is the protein complex of the nuclear pore, or the pore complex (Figure 2.17), which is formed by different proteins with a total molecular weight up to 1200 kDa. Among these proteins, the presence of ATPase and acto-myosin complexes has been demonstrated.
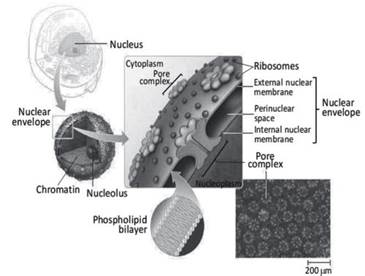
Figure 2.17. Diagram of the pore complex in a nucleus of a generic eukaryotic cell. The diagram in the center shows the structure of the nuclear envelope. In the inserts are electron microscopic images of the phospholipid bilayer and the surface of the nuclear envelope, showing the density and spatial distribution of the pore complexes
The pore complex is anchored to the introflexions that the nuclear envelope presents for the purpose of allowing insertion of the pore. Therefore, the discontinuities of the nuclear envelope at the pores do not correspond to an interruption of either the external or internal membrane, but only to the folds that ensure the continuity of the double phospholipid layer (Figure 2.16).
Under physiological conditions, nucleocytoplasmic communication is guaranteed by a pore opening of about 9 nm, which allows free diffusion of ions and small solutes up to a molecular weight of 40 kDa. The pore complex appears to be mainly involved in the transport of RNA from the nucleus to the cytoplasm, although the molecular mechanism has not yet been clarified. The use of a peptide, WGA (Wheat Germ Agglutinin), that is able to interact with the pore complex interrupts the passage of RNA.
The other membranous complex presenting a double phospholipid layer is the mitochondria (Figure 2.18). Many scholars believe that mitochondria were originally autonomous bacterial organisms that later became symbionts with cells during evolution. Their main function is to produce ATP through the Krebs cycle using glucose.
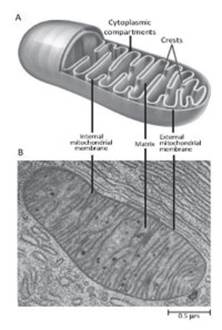
Figure 2.18. Structural scheme (A) and electron microscope ultrastructure (B) of a generic mitochondrion
The enzymes of the respiratory chain are supported by the internal mitochondrial membrane, which folds into the characteristic "ridges" and increases the useful surface area. The outer membrane has several permeation pathways through the single phospholipid bilayer. There are also proteins that pass through both membranes at the points where they pack together, areas called contact sites.
These proteins are real ionic channels called VDAC, or Voltage-Dependent Anion Channels, and ensure the modulation of solute exchanges between the cytoplasm and mitochondrial matrix. The importance of these permeation pathways is manifested in the enormous potential energy that the mitochondria are able to accumulate, in the form of an electrical potential difference between the two membranes that reaches values close to -200 mV (Figure 2.19) and which guarantees sufficient energy for the production of ATP.
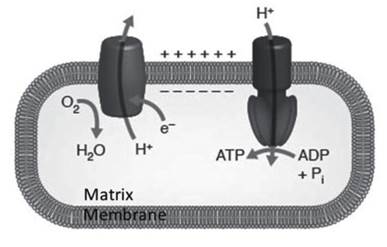
Figure 2.19. Scheme of the exploitation of the mitochondrial potential difference for ATP production
There are also several other membranous formations inside the cell, such as the endoplasmic reticulum, which can have many specializations in cells with different functions. In particular, the ability of the endoplasmic reticulum to accumulate calcium ions to be used as an intracellular messenger will be considered.
The Golgi apparatus, lysosomes and endosomes are just some of the many other intracellular structures that use phospholipid membranes to isolate environments within the cell from the rest of the cytoplasm, in order to diversify the functions that are required in an adult cell to ensure its proper function.
Date added: 2024-07-02; views: 592;