Stress and Growth Regulation
Growth is an increase in biomass, volume, area or length as a result of highly coordinated cell division, cell expansion and cell differentiation. Stem cell populations are maintained in meristems. They produce new cells, which then expand and acquire a fate, that is, a specific role within a tissue and organ. All of these processes are under hormonal control and are responsive to both internal and external factors.
The integration of environmental cues into a growth response is the basis for a hallmark of plant biology: the plasticity of developmental steps and organ formation (Pierik and Testerink 2014). In contrast to animals, which are born with a complete set of organs, plants develop their organs only after embryogenesis and germination. The morphology of a plant individual not only is the realisation of a blueprint but also arises under strong influence from environmental factors. Light conditions govern leaf morphology, and nutrient availability in the soil determines root architecture. Charles Darwin studied phototropism, the growth of a plant towards a light source (Darwin 1880). These and many other examples illustrate that the regulation of growth and development is an integral part of stress physiology (for more details on plant development, see plant physiology and plant molecular biology textbooks).
The strain caused by a stress factor could be regarded as a perturbation of the complex processes that constitute development, necessarily causing stagnation of growth. Indeed, growth reduction is widely seen and interpreted as an indicator of stress. Such effects can be defined as passive growth reduction due to interference of stress factors, such as water scarcity, with metabolic and physiological processes that are important for growth. However, there is also acclimation via active growth modulation. A plant must always balance resource allocation between investment in growth and stress acclimation (Fig. 2.24). In the past decade, molecular insights into the mechanisms of hormonal growth control have provided direct evidence of this fundamental trade-off.
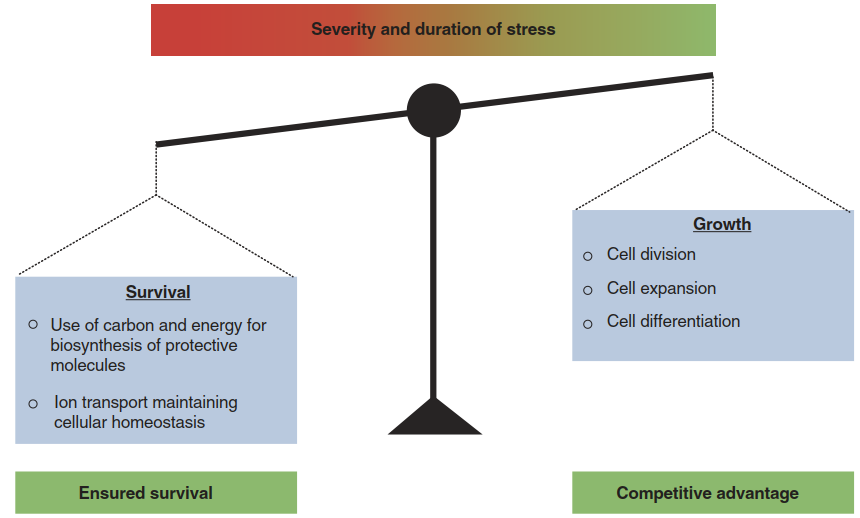
Fig. 2.24. The fundamental trade-off between growth and stress acclimation. Plants need to find the right balance in resource allocation between growth and survival depending on the severity of stress
Gibberellic acid (GA) is one of the classic plant growth hormones. Among other effects it promotes the elongation of plant organs such as internodes. The so-called DELLA proteins, which are negative regulators of the GA response, are key components of active growth modulation in plants (Santner and Estelle 2009). DELLA proteins repress the transcription of GA-responsive genes. The soluble GA receptor protein GID1 associates more strongly with DELLA proteins when GA is bound. This interaction promotes the tagging of DELLA proteins for degradation by the 26S proteasome and thereby the release of a GA response brake (Fig. 2.25). When A. thaliana mutants lacking most of the DELLA proteins are exposed to salt stress, they do not show the same reduction of root growth that is displayed by wild- type seedlings (Achard et al. 2006).
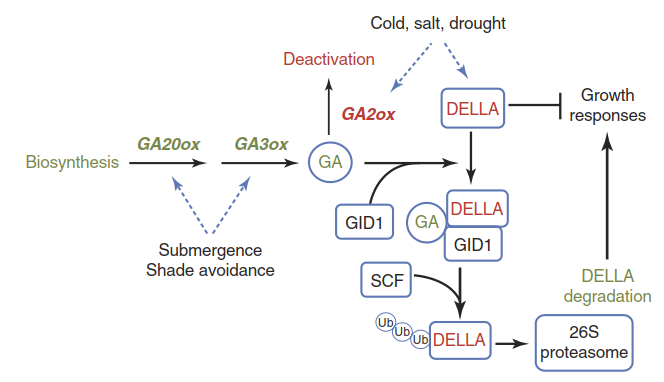
Fig. 2.25. The gibberellic acid (GA) signalling pathway and DELLA proteins are key regulators of growth under stress conditions. Stresses such as cold, salt and drought trigger active growth reduction, while acclimation to shade and submergence often involves growth stimulation. GA20ox (GA20 oxidase) and GA3ox are GA biosynthesis enzymes; GA2ox is an enzyme catalysing a step in GA turnover. GID1 is the soluble GA receptor. Upon GA binding it associates with an SCF E3 ligase, which tags DELLA proteins for degradation by poly-ubiquitination
This simple experiment clearly demonstrates that growth is actively slowed down under stress and in a manner dependent on DELLA proteins. Salt stress does not per se inhibit growth under the tested conditions. The consequence for the mutant seedlings is a lower survival rate. They maintain growth at the expense of an acclimative response, such as osmotic adjustment, and eventually die. Similar outcomes can be observed under drought conditions. Plants with an artificially high GA status (e.g. through overexpression of GA biosynthesis genes) grow and compromise survival, while plants with a low GA status (e.g. DELLA mutants) remain small and survive (Colebrook et al. 2014). Under natural conditions they would of course suffer a disadvantage in competition with neighbouring plants (Fig. 2.24).
Not all stress conditions trigger active growth reduction. Plants often attempt to escape a low-light condition caused by other vegetation through a shade avoidance response, that is, an investment in more growth, especially of the stem and the petioles. Some species adapted to flooding-prone habitats try to reach the water surface when they are submerged. This strategy is to ensure continuing oxygen supply to tissues under water. As in the case of shade avoidance, the up-regulation of GA biosynthesis is a component of this strategy (Fig. 2.25).
Date added: 2025-01-13; views: 545;