Photoperiodism. Control of Flowering by the Photoperiod
How does a plant determine that the right day length for the initiation of flowering in spring or for cessation of growth and bud set in autumn has arrived? Photoreceptor-mediated perception of light is a prerequisite but is not sufficient, because no information on the duration of light exposure is recorded. The mechanisms enabling the measurement of the day length have been elucidated in A. thaliana and in the monocot model rice. A. thaliana is a facultative long-day plant— that is, long days trigger inflorescence development—but even without this cue, flowering will eventually happen. The same mechanisms, however, also explain the behaviour of short-day plants and even the basis of other photoperiodic processes such as bud set in aspen trees. This illustrates the value of a model system, as discussed in Sect. 2.2.5.
Classic grafting experiments led to the concept of florigen, a flowering hormone that remained hypothetical for decades. Florigen refers to the chemical signal that travels from leaves to the shoot apical meristem to deliver the information that the right day length has arrived for the switch from vegetative to reproductive growth. When one leaf of a plant (e.g. Perilla frutescens var. crispa, common name Shiso, Lamiaceae) is exposed to the right day length for flowering and this leaf is grafted onto a plant not exposed to the right day length, flowering is induced nonetheless. Such a transmission of information via grafting is even possible between related species—for example, the Solanaceae Petunia and tobacco (Zeevaart 2006).
After many years of futile attempts to identify florigen, the careful analysis of A. thaliana flowering mutants (Koornneef et al. 1991) finally revealed the core photoperiodic mechanism (Yanovsky and Kay 2002). It depends on the circadian rhythm in the abundance of CONSTANS (CO) transcript. CO encodes a key regulator of flowering. The transcript level peaks late in the afternoon (Fig. 2.32). However, CO protein accumulates only when light is still present at the time of the highest transcription. Light is perceived by the photoreceptors cryptochrome and phytochrome. In the dark the CO protein is very unstable and quickly degraded.
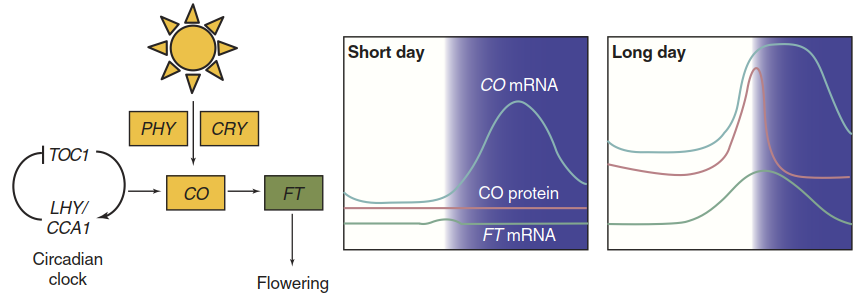
Fig. 2.32. Control of flowering by the photoperiod. The day length is perceived via the coincidence of an internal factor and an external stimulus. The central regulator CONSTANS (CO) is under control of the circadian clock (TOC1 and LHY/CCA1 are key components of the clock). Under short-day conditions the peak in transcript abundance occurs in the dark and the protein does not accumulate. It is only when light is perceived by the photoreceptors phytochrome (PHY) and cryptochrome (CRY) at the time of high CO expression that CO protein accumulates and activates the synthesis of the FT protein, florigen. (Modified from Buchanan et al. (2015))
Thus, CO is active only upon the coincidence of two events: the regularly recurring peak in transcription controlled by the intrinsic biological clock, and exposure to the external factor light (external coincidence model; its history was described by Kobayashi and Weigel 2007). For A. thaliana this happens only when the day length exceeds a certain threshold, that is, when the sun sets after the time of maximal CO transcript abundance. Active CO protein then triggers the synthesis of FLOWERING LOCUS T (FT) in the leaf. The FT protein is florigen. It travels via the phloem to the shoot apical meristem and signals the right day length for flower induction (Fig. 2.33).
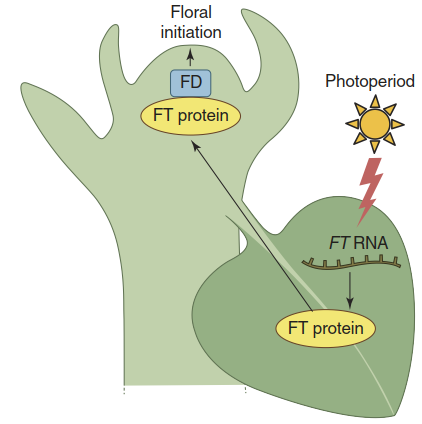
Fig. 2.33. Induction of flowering by the FT protein, florigen. It is synthesised in leaves under control of the photoperiod (Fig. 2.32) and travels via the phloem to the shoot apical meristem, where it heterodimerises with FD, a bZIP transcription factor, and initiates the formation of flowers (Buchanan et al. 2015)
The module consisting of CO and FT is essential for other photoperiodic processes as well, and is likely to explain such phenomena throughout the plant kingdom. An analysis of aspen trees growing at different northern latitudes demonstrated a key role of CO and FT in local adaptation (Fig. 2.34). Aspen trees enter a state of dormancy at the end of summer. Growth ceases, buds are set and cold tolerance increases (Fig. 2.26).
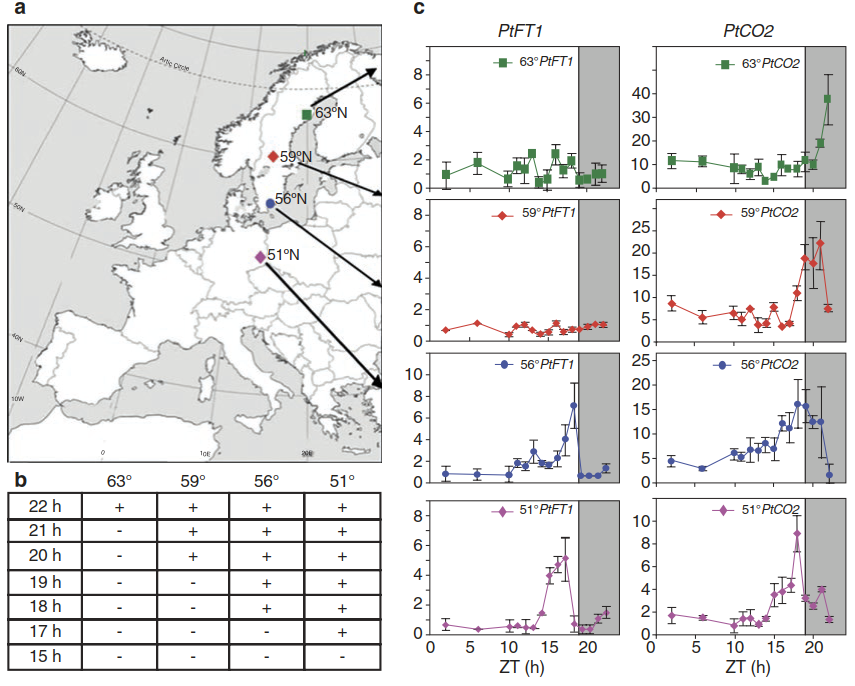
Fig. 2.34. Variation in the photoperiodic control of transition to dormancy in aspen trees from different northern latitudes. a Map of Europe indicating the origin of the four different European aspen (Populus tremula) clones. The green squares indicate Umea (63 °N), the red diamonds indicate Brunsberg (59°N), the blue circles indicate Ronneby (56°N) and the lilac diamonds indicate Brauna (51°N). b Critical day lengths for growth cessation in the collected P tremula clones; + indicates growth and — indicates bud set. c PtFTI and PtCO2 expression in the different P tremula clones in a 19-h-light/5-h-dark cycle. The shaded boxes indicate night and the white boxes indicate light (Bohlenius et al. 2006)
These processes are triggered when the day length falls below a critical threshold. In northern Sweden, winterly conditions arrive earlier than in Germany, for example. Thus, aspen trees need to go through this transition into dormancy at a time of the year when the days are still comparatively long, while in Germany they maintain growth until the day length is already much shorter (Fig. 2.34b). An important aspect of local adaptation to the stress conditions of a cold winter is therefore to respond to the day length change that matches the specific seasonal pattern of the habitat. FT in Populus tremula suppresses the short-day-induced cessation of growth and bud set, that is, as long as FT reaches the meristems, they continue to grow (Bohlenius et al. 2006).
When the transcript abundance of CO and FT in P tremula individuals originating from different latitudes was analysed in identical 19-h-light-5-h-dark cycles, it was found that the endogenous rhythm of CO transcript accumulation was different. In high-latitude individuals the peak of CO is shifted to later in the day and reaches its peak only after the end of the light period in a 19-h-light/5-h-dark cycle. As a consequence, FT expression is no longer activated, the signal supporting growth is absent and a state of dormancy is initiated. In contrast, the CO peak in individuals from lower latitudes reaches a peak earlier in the day. Hence, FT is expressed and growth is maintained in a 19-h day.
Such observations demonstrate that local adaptation to the pattern of seasonal changes can occur via the fine-tuning of endogenous oscillations in the expression of central regulators such as CO. Moreover, the challenges arising from rapid climate changes, especially for perennial plants such as aspen trees, become obvious. When seasonal changes in temperature or precipitation can no longer be reliably anticipated by measuring the day length, organisms will suffer from a loss of fitness. They might initiate dormancy too early (thereby reducing biomass production) or too late, hence running a greater risk of damage through unfavourable conditions.
Date added: 2025-01-13; views: 603;