Developmental Switches Triggered by Favourable Conditions
The preceding subchapters described molecular mechanisms enabling a plant to switch to reproduction at the right time by anticipating favourable conditions. Another developmental transition during a plant’s life cycle, which is of at least equal importance for adaptation, is seed germination (Donohue et al. 2010). It determines when a new plant individual enters an ecosystem (Bewley 1997). A mature seed in its dormant state is normally well protected from abiotic and biotic stress factors. For example, its low water content practically rules out pathogen infection. Water scarcity is not a problem and the seed coat protects against damaging doses of radiation.
When a seed germinates it becomes much more vulnerable to stress factors. Thus, undergoing this dramatic transition under the wrong circumstances entails an acute risk of death. The appropriate timing of germination is therefore an important aspect of an escape, possibly illustrated best by desert blooms. Many desert-adapted plant species are actually not able to cope with the typical conditions of a desert: scorching heat and lack of water. Still, they survive in such habitats because their seeds can lie dormant in the soil for many years and germinate only after one of the rare rain events allows a few weeks of vigorous physiological activity (Fig. 2.36).

Fig. 2.36. Desert bloom in the Atacama Desert
What is critical for a successful escape is germination under favourable conditions. The ability to germinate is dependent on the dormancy of a seed, which imposes a delay between the shedding of a seed by the mother plant and the germination of the seed. A dormant seed is not able to germinate even when conditions such as water and permissive temperatures are present that would physiologically allow germination in the non-dormant state (Finch-Savage and Leubner-Metzger 2006). Thus, dormancy defines the conditions under which germination occurs, and dormancy has usually evolved in such a way that a seed of a particular plant germinates under environmental conditions that are expected to favour seedling establishment and survival.
Many forms of dormancy are known— for example, physical dormancy, caused by a water-impermeable seed coat. Most widespread is physiological dormancy, and mechanistic understanding of this is available for only a few species. One of the molecular factors involved in establishing a state of dormancy is ABA. It functions as a positive regulator of dormancy, as shown by the early germination of many ABA biosynthesis and ABA perception mutants. The ABA antagonist is GA. This growth hormone can break the dormant state, and GA biosynthesis mutants of A. thaliana are unable to germinate (Finkelstein et al. 2008). A simplified view of dormancy regards it as a state that is controlled by the balance—that is, the ratio of concentrations—between the two hormones.
In accordance with the hormone balance concept, release of dormancy is associated with a shift in the ABA to GA ratio towards lower values through GA biosynthesis and ABA breakdown. Mature seeds continuously perceive information on environmental conditions. This information is used to decide on the right time for germination. Several different environmental factors can trigger the release of dormancy, and there is tremendous variation between plant species as to which factors are decisive (Lambers et al. 2008) (Fig. 2.37). A very common factor is a stimulating effect of light on seed germination. Light exposure can indicate the absence of competing vegetation above, which would give the developing seedling the chance to escape low-light stress.
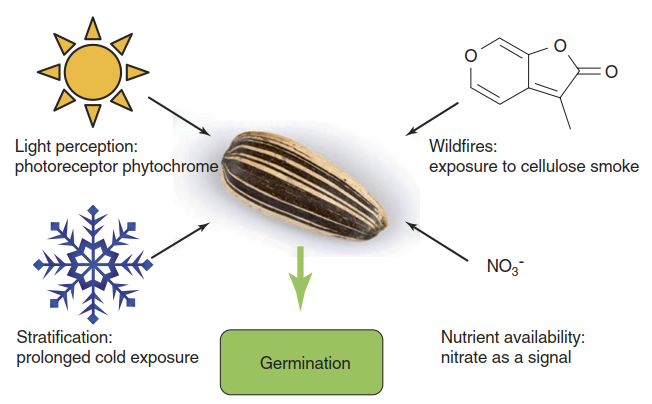
Fig. 2.37. Triggers of seed germination. Diverse environmental stimuli can be used as indicators of favourable conditions
Also, the light requirement prevents germination of a seed buried too deeply in the soil to reach the surface. It has been known for a long time that the red light receptor phytochrome (Chap. 3) mediates this response. Red light absorption by the chromophore triggers a conformational change in phytochrome to its physiologically active far-red-absorbing form, Pfr. A high far-red to red ratio, which is typical for shade caused by vegetation, therefore inhibits the response (Chap. 3). Active phytochrome represses ABA biosynthesis and stimulates GA biosynthesis. Furthermore, it interacts with repressors of germination such as PIL5 (phytochrome interacting factor 3-like5) and causes their degradation (Holdsworth et al. 2008).
Many plants in seasonally changing climates produce seeds that should germinate only after the winter is over. For example, seeds of summer annuals shed in autumn require extended exposure to cold. This process is termed stratification and applies to imbibed seeds, that is, seeds that have taken up water and are now waiting for the right conditions to proceed to germination. Stratification activates GA biosynthesis. How the winter memory functions molecularly is not understood but may well be based on processes similar to the ones unravelled for vernalisation.
Nitrate and molecules contained in smoke are chemical germination stimulants. Nitrate not only is an important nutrient for plants but also can serve as an indicator of nutrient availability and the absence of competing vegetation. As this is typical for a disturbed site, seeds of ruderal species particularly respond to nitrate. How nitrate sensing is translated into a modulation of the ABA to GA ratio is molecularly unresolved.
Wildfires occur in many habitats around the world and are frequent especially in hot and dry Mediterranean-type climates. After a fire the chances of successful seedling establishment are often strongly improved because shading vegetation cover is removed and nutrient availability may be greater. Several plants are so highly adapted to such conditions that germination of their seeds is strictly dependent on fire. It can proceed only after compounds released by the combustion of plant biomass are perceived. Seeds of many plant species, even those of A. thaliana and others that normally do not grow in fire-prone habitats, are stimulated to germinate by smoke exposure (Nelson et al. 2012).
Burning cellulose and hemicellulose are very good sources of volatile germination stimulants. The first compound to be isolated was a butenolide (Flematti et al. 2004) (Fig. 2.37). It was found to be active at concentrations below 1 part per billion (ppb). Later, several closely related molecules were isolated and they are now collectively called karrikins. Interestingly, these karrikins are very similar to strigolactones, which are recently discovered plant hormones and stimulants of seed germination for parasitic plants (Chap. 8). At least one molecular factor (called MAX2, for MORE AXILLIARY GROWTH2) is required for the responses to both strigolactones and karrikins (Nelson et al. 2012). Thus, the signalling pathways most likely overlap.
Date added: 2025-01-13; views: 571;