Avoidance of Ultraviolet-B- Induced Stress
Plants synthesise a range of compounds in order to avoid damage by UV-B radiation. The rate of accumulation in different plant types correlates well with the average UV exposure associated with the respective habitat (Fig. 3.23).
Phenylpropanes, as aromatic compounds, exhibit strong absorbance of shortwave radiation and are thus able to function as effective UV filters or sunscreen pigments. Many compounds in this family of secondary plant metabolites absorb light only in the UV range and cannot be recognised as pigments by the human eye. Others, such as the anthocyanins, absorb light also in the visible range and therefore appear as pigments (Figs. 3.25 and 3.26).

Fig. 3.25. Anthocyanins in flushing leaves of the tropical Sapotacea Inhambanella henriquesii in the coastal forests of Kenya. The anthocyanin is located in the vacuoles of the leaf epidermis and protects the leaves, which are not yet fully green, from radiation damage. The close-up (right) shows the mixture of colours between the red anthocyanins and the chlorophyll. (Photos: E. Beck)
Glycosylation renders phenylpropanes water soluble so they can be sequestered in the vacuoles primarily of the epidermis. In certain purple varieties such as copper beech (Fagus sylvatica), the vacuoles of the mesophyll cells contain such pigments too. They absorb in the UV spectrum (Fig. 3.26) and in the green region of the visible spectrum and thus do not interfere with the absorption of light by the chlorophylls. Protective colouration of the leaves by anthocyanins is frequently observed in unfolding young leaves (called “juvenile anthocyanin”) and again during senescence.
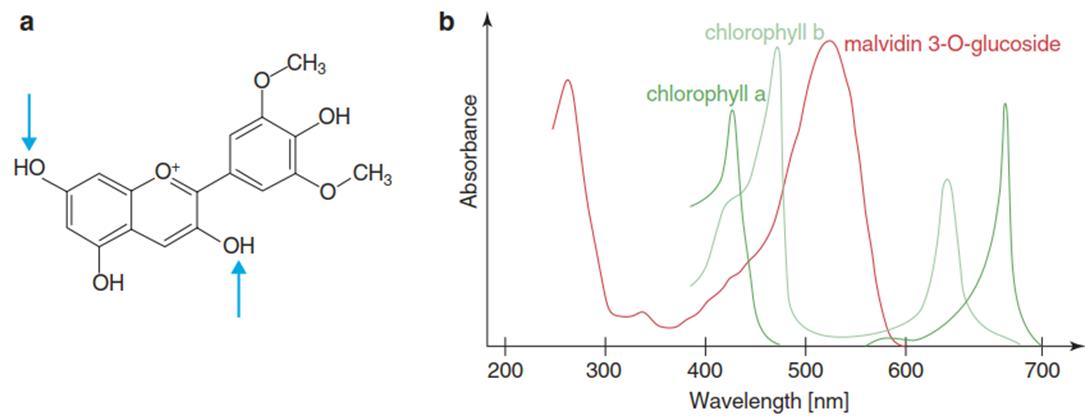
Fig. 3.26. After glycosylation, phenylpropanes are accumulated in the vacuoles of the epidermal cells. a Chemical structure of the anthocyanidin malvidin (at acidic pH, arrows indicate the positions for glycosylation) and b the absorption spectrum of its 3-O-glucoside (which is nearly identical to that of the aglycone malvidin) in comparison with the absorption spectra of chlorophylls a and b
In young leaves the additional pigments protect the developing photosynthetic machinery until the energy dissipation mechanisms are fully functional. The “autumn anthocyanin” is well known from the Indian-summer aspect of deciduous trees. Its ecological function, however, is not well understood. Most likely it protects the controlled degradation of leaf constituents—in particular, the chlorophylls—and the export of degradation products to the overwintering parts of the plant. The key enzyme in the flavonoid metabolism is chalcone synthase, whose formation is strongly induced by UV-B as well as by UV-A/blue light.
Other examples of compounds—out of the great variety of aromatic secondary metabolites— that accumulate upon UV-B exposure include 3",6"-DCA and 3",6"-DCI (=3" ,6" -di-para-coumaroyl-astragalin and 3",6"-di-para-coumaroyl-isoquercitrin) in pine needles and 2" ,2"-di-para-coumaroylkaemperol-3α-d- arabinoside in beech leaves (Figs. 3.27 and 3.28).
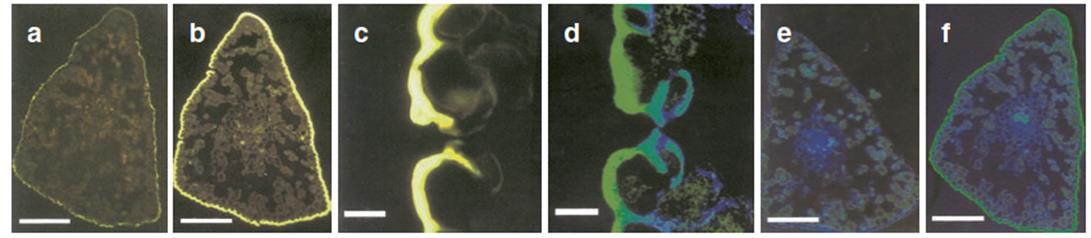
Fig. 3.27. Histochemical evidence for ultraviolet (UV)-absorbing substances in the epidermis of pine needles. a-c Fluorescence (excited by light of 450-490 nm wavelength, determined at >520 nm): a control; b, c after treatment with a reagent that intensifies flavonoid fluorescence. d-f Observation with a confocal laser microscope (which shows the cells and their content more clearly): (e) control; d, f after staining, as above. a, b, e, f Transverse section of a needle; c, d stomata. (Modified from Schnitzler et al. (1996))
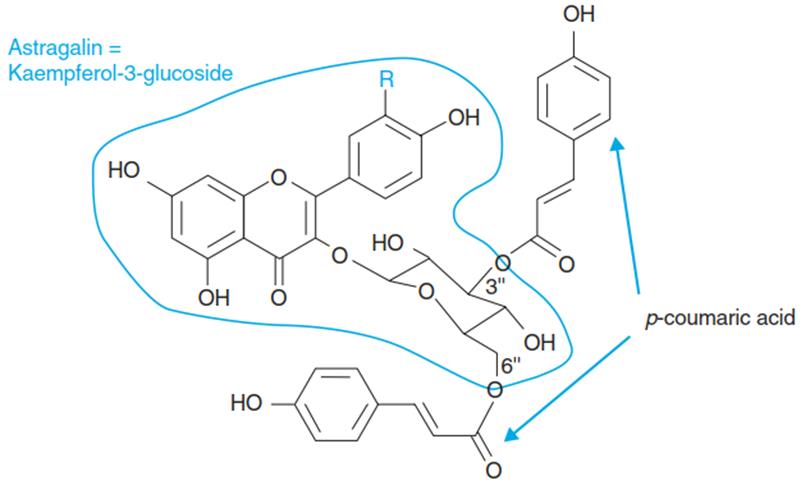
The non-conjugated phenylpropanes (e.g. p-coumaric acid and its derivatives) are primarily localised in the epidermal cell wall, while the conjugates are mainly sequestered in the vacuoles of the same cells. It has been calculated that the accumulation of aromatic compounds in the outer epidermis cell wall of pine needles allows only about 4% of the incoming UV-B to pass through. Diacylated flavonoids in the vacuole are able to filter out the residual UV-B, leaving the mesophyll completely unaffected. This explains the exceptional UV protection that needles of coniferous trees are known for (Fig. 3.23).
The association between concentrations of UV screens and UV exposure in the natural habitat is apparent when comparing, for instance, alpine herbs with herbs from the lowlands (Filella and Penuelas 1999; Fig. 3.23). Moreover, because synthesis of UV-B screening phenols and phenylpropanes is stimulated by UV-B radiation, variations in their concentrations can be expected also when leaves of the same species but from locations with contrasting UV-B radiation intensities are compared.
One example of such a local adaptation is mountain avens (Dryas octopetala) from the Arctic, from southern Norway and from the French Alps, where the average UV-B radiation is three times higher than in the Arctic. UV-B transmittance in the epidermis of Dryas from the French Alps was only 2.5% versus 5% in plants from Norway and 7% in Arctic plants (Nybakken et al. 2004). It was also shown that depending on the weather conditions during the year, the screening capacities of the leaf epidermis may change, indicating a dynamic acclimation response (Barnes et al. 1996).
Date added: 2025-01-17; views: 505;