Signal Transduction: The Signalling Centre COP1
Growth of plants in the dark, known as etiolation or skotomorphogenesis, requires suppression of photomorphogenesis, which is initiated by illumination. Activation of the above described photoreceptors - phytochromes and cryptochromes in particular - by irradiation triggers signal transduction cascades, which converge at a protein termed COP1 (CONSTITUTIVE PHOTOMORPHOGENIC 1).
COP1 is a central switch which, because of its complex structure, can interact with several other proteins. In the dark, COP1—by its function as an E3 ubiquitin ligase—targets photomorphogenesis-promoting transcription factors for degradation by the ubiquitin-proteasome system, thus preventing photomorphogenesis.
One of these proteins is the transcription factor ELONGATED HYPOCOTYL 5 (HY5) which, together with others, is responsible for the onset of photomorphogenesis (e.g. the inhibition of hypocotyl elongation growth). In the dark, it is degraded in the proteasome, giving rise to the well-known etiolated hypocotyls. Light-activated photoreceptors inactivate COP1 by mediating its export from the nucleus, thereby suspending its activity against transcription factors that are located in the nucleus.
For a further understanding of the interaction of COP1 with the photoreceptors, it is necessary to introduce another protein, SPA1 (SUPPRESSOR OF PHY A 1), which forms a complex with COP1 to enable its association with the E3 ligase core complex; this finally ubiquitinates the proteins, targeting them for degradation (Fig. 3.21a). In the light, activated Cry1 and 2 interact with SPA1, dissociating it from COP1, which can now be exported from the nucleus (Fig. 3.21b). Further targets of the COP1 E3 ubiquitin ligase are the phytochromes—in particular PhyAfr and cryptochrome 2.
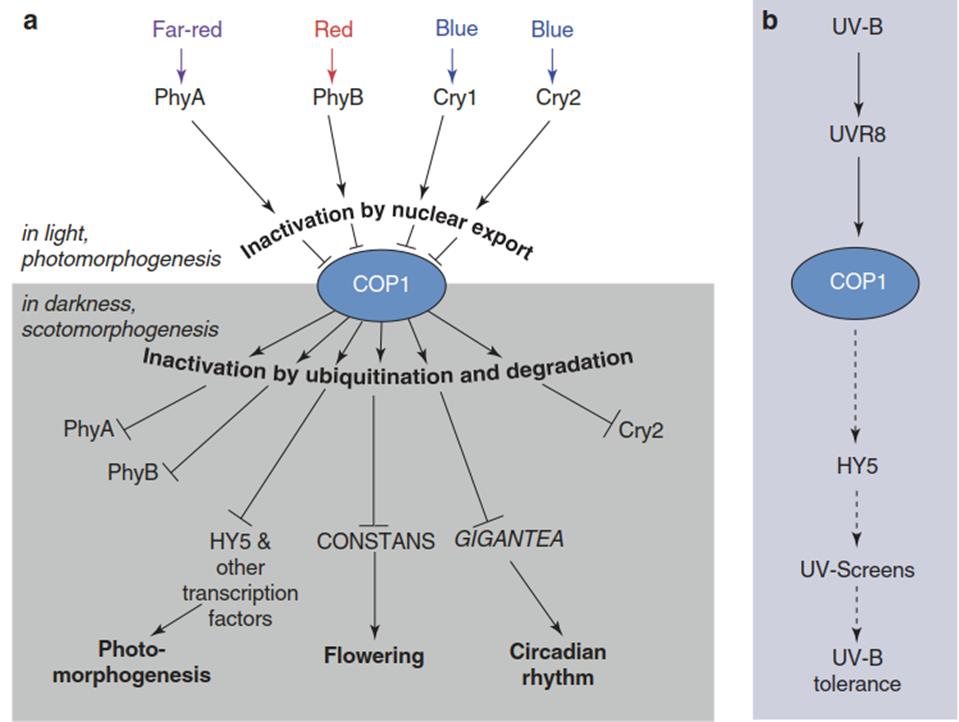
Fig. 3.22. Roles of the central regulator COP1 in signal transduction of skotomorphogenesis, photomorphogenesis a and ultraviolet (UV)-triggered responses b. In addition to photomorphogenesis, COP1 is involved in other developmental events (regulation of flowering, with the transcriptional regulator CO (CONSTANS)) and the circadian rhythm (by regulating the abundance of the circadian-associated protein GI (GIGANTEA)). (Modified from Lau and Deng (2012))
While COP1 is a negative regulator of photomorphogenesis, it promotes UV-B-triggered reactions of plants, hence acting as a positive regulator (Fig. 3.22). To understand this difference, it is important to note that in skotomorpho- genesis, COP1 acts as a homodimer, and also the COP1-SPA1 complex is dimeric. Dissociation of the COP1-SPA1 complex by sequestration of SPA1 through interaction with cryptochrome also monomerises the COP1 homodimer. In the cytosol, monomeric COP1 can bind to the monomeric activated UVR8 receptor and the heterodimer migrates back into the nucleus, where it activates HY5 and other transcription factors and effectors.
Date added: 2025-01-17; views: 594;