Temperature and Photosynthesis
Practically any type of stress that impairs CO2 assimilation in the Calvin cycle under high light can cause oxidative damage and photoinhibition (of photosynthesis). Low temperatures are among the most prominent of these stresses. Essentially, two processes account for oxidative damage. First, excitation energy cannot be channelled efficiently into the reduction of nicotinamide adenine dinucleotide phosphate (NADP+), because the balance is disturbed between photosynthetic thylakoid reactions (the so-called light reactions, which are virtually not decelerated), and the biochemical CO2 fixation reactions which, in contrast, are slowed down considerably in the cold. As a consequence, reduced NADP (NADPH) is not oxidised at a sufficient rate in the reducing phase of the Calvin cycle, leading to a shortage of the electron acceptor NADP+.
Under such conditions, electrons are transferred to O2 instead and thereby radicals and reactive oxygen species (ROS) are produced. Depending on the scavenging capacity of the cell, ROS can be detoxified or cause damage. In addition, they can act as signals activating pathways that target genes for ROS-detoxifying proteins such as ascorbate peroxidase, glutathione reductase or superoxide dismutase.
ROS generation can inhibit the repair of the D1 protein, which is the primary site of photoinhibition due to excessive light intensity (Yamamoto 2016). The D1 protein is a component of photosystem II (PS II), which contains—in addition to the paired P680 reaction centre—several other photopigments, as well as the quinones QA and the mobile QB (plastoqui-none in the bound state). During unlimited electron drainage from PS II to plastoquinone the excited chlorophylls of P680 are in the singlet state 1P680, which readily dissipates its energy through ChlDI and pheophytinD1 to QA.
Impaired reoxidation of QAθ — e.g. because of over-reduction of plastoquinone or other components of the linear photosynthetic electron transport—results in a spin conversion of P680+QAθ to 3[P680+QAθ] (triplet state). This triplet state of P680 reacts readily with molecular oxygen 3O2, producing the extremely reactive singlet oxygen 1O2. (Quenching of singlet oxygen formed at a low rate by carotenoids is described in detail in Chap. 2, Sect. 2.2.3). At low temperatures, quenching cannot keep pace with the production of singlet oxygen.
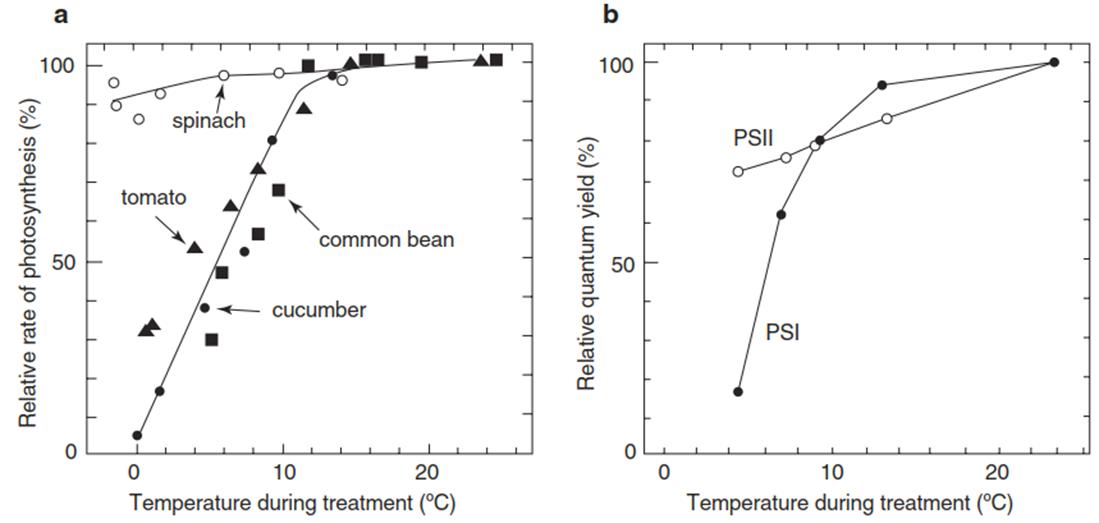
Fig. 4.3. Photoinhibition by chilling at a photon flux density (PFD) of about 450 pmol quanta m-2 s-1 upon exposure for 2.5 h to the indicated temperatures. The rate of photosynthesis by leaf slices maintained at 25 °C at the same PFD is 100%. a Photosynthetic CO2 net uptake. b Photosynthetic quantum yield of photosystem II (H2O → 2,6-dimethyl-p-quinone) and photosystem I (diaminodurene → methylviologen/O2), respectively, measured in potato leaves. Cucumber, common bean, tomato and potato are chilling sensitive; spinach is chilling tolerant (Sonoike 1996)
This is the second process (besides ROS production) that causes photoinhibition of PS II. A similar process has also been described for photosystem I (PS I), albeit with ROS species other than singlet oxygen. Here the equivalent to the D1 protein, PsaB, is damaged when ROS production (in particular, the hydroxyl radical) exceeds detoxification. Photoinhibition of PS I is particularly pronounced in tropical species sensitive to chilling—that is, cold temperatures above freezing (Fig. 4.3). The difference in the damage to D1 and PsaB is the time required for repair. Recovery from photoinhibition of PS II requires only hours, whereas restoration of PS I takes several days.
Date added: 2025-01-17; views: 540;