Variations in Temperature Range
A classification most commonly used in microbiology sorts organisms into three categories based on their temperature optima: psychrophiles, mesophiles and thermophiles. Psychrophiles are organisms adapted to cold habitats where water frequently changes from the liquid to the crystalline state. Most of them are unicellular snow and ice algae (e.g. Haematococcus pluvialis and various diatoms) or bacteria that prevent freezing of cellular water. At the other end of the scale are the thermophiles (and extremophiles) with optimal temperatures for growth between +80 °C and above 100 °C.
They comprise certain unicellular algae such as Cyanidioschyzon merolae, cyanobacteria, bacteria and archaea, most of them living in hot springs, in geysers or on the black smokers of the deep sea. Most organisms, including practically all plants—in particular, the (homoiohydric) terrestrial and the marine species—belong to the meso- philes, which thrive at day temperatures between 15 °C and 25 °C and (in the case of terrestrial species) night temperatures about 10 K lower.
However, while temperature optima are not that different between plants, the tolerable temperature range varies widely (Tables 4.3 and 4.4). Most of the variation is attributable to differences in the ability to withstand temperatures below the optimum. The temperature at which cold damage occurs largely corresponds to the geographical distribution of a species. For instance, plants from temperate zones are usually more cold tolerant than plants from tropical regions. They are able to withstand sub-zero temperatures, whereas many tropical plants already show damage when exposed to temperatures slightly above freezing (chilling) (Fig. 4.3).
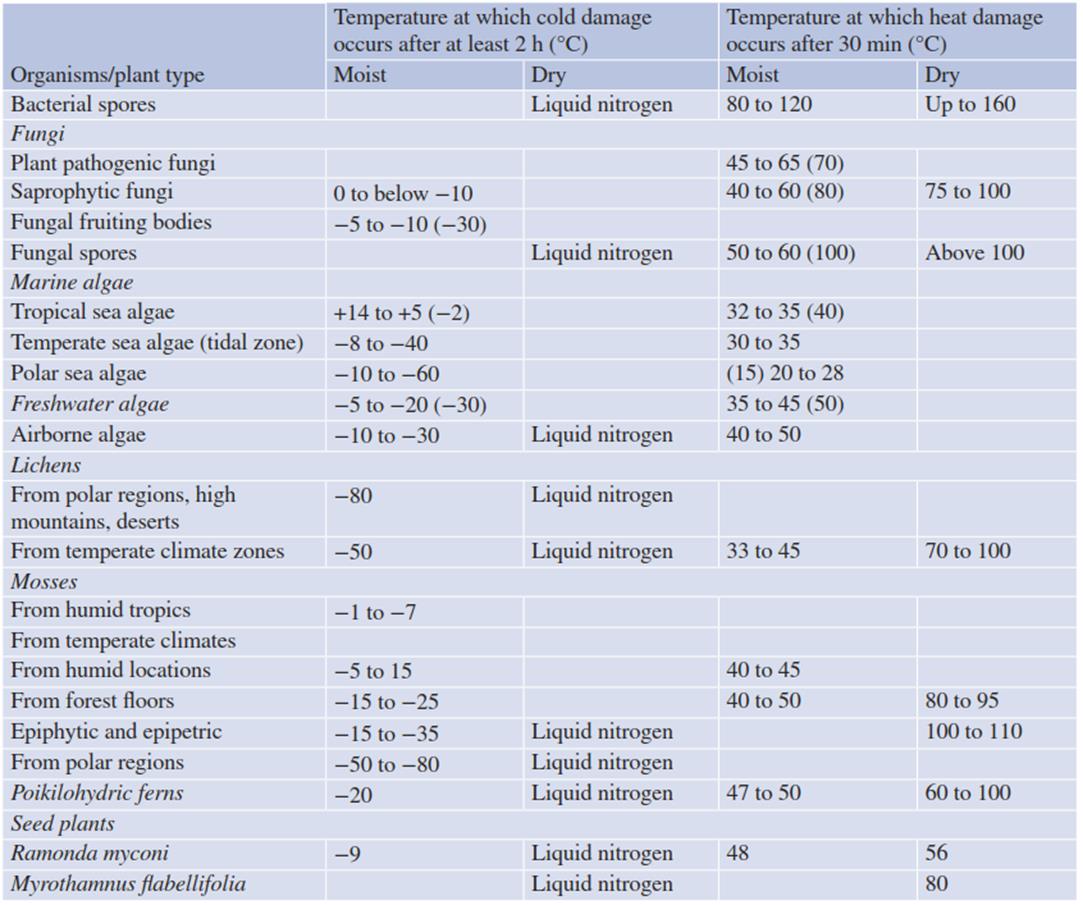
Table 4.3. Maximum temperature resistance of microorganisms and poikilohydric seed plants in the moist turgescent state and in the dry rigid state caused by desiccation
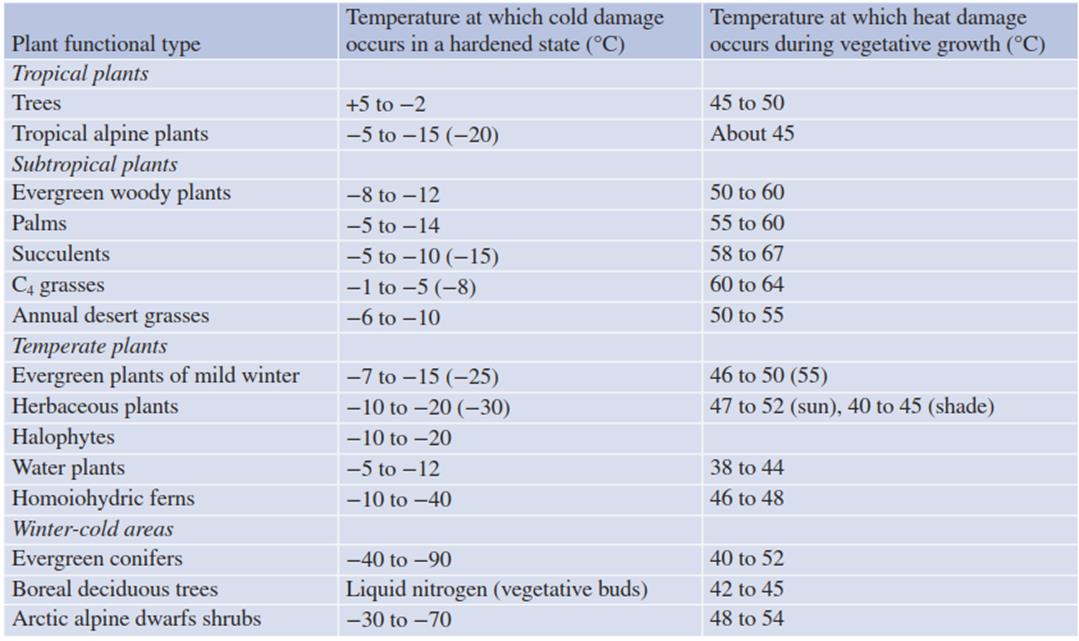
Table 4.4. Temperature tolerance of the leaves of vascular plants from various climate zones
Palms, mangroves, and tropical ornamental plants such as the African violet (Saintpaulia ionantha) (Fig. 4.8) or the coffee tree (Fig. 4.9) have been studied intensively. They are all adapted to a temperature range of between +5 and about 50 °C. Many other crops are chilling sensitive as well (e.g. cotton, soybean, maize and rice).
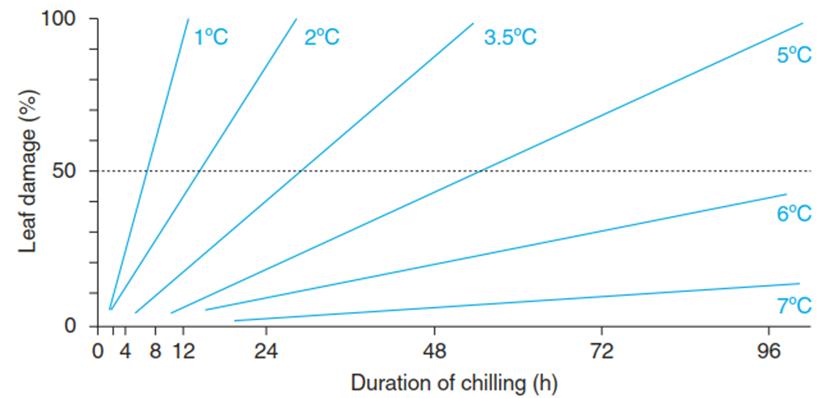
Fig. 4.8. Damage to the African violet (Saintpaulia ionantha) by various chilling treatments. Below the threshold temperature of +8 °C, necrosis occurs. The strength of the chilling stress can be estimated as the product of cold (above the freezing point) multiplied by the duration of the exposure to the temperatures, as indicated in the figure. (Modified from Larcher (1994))
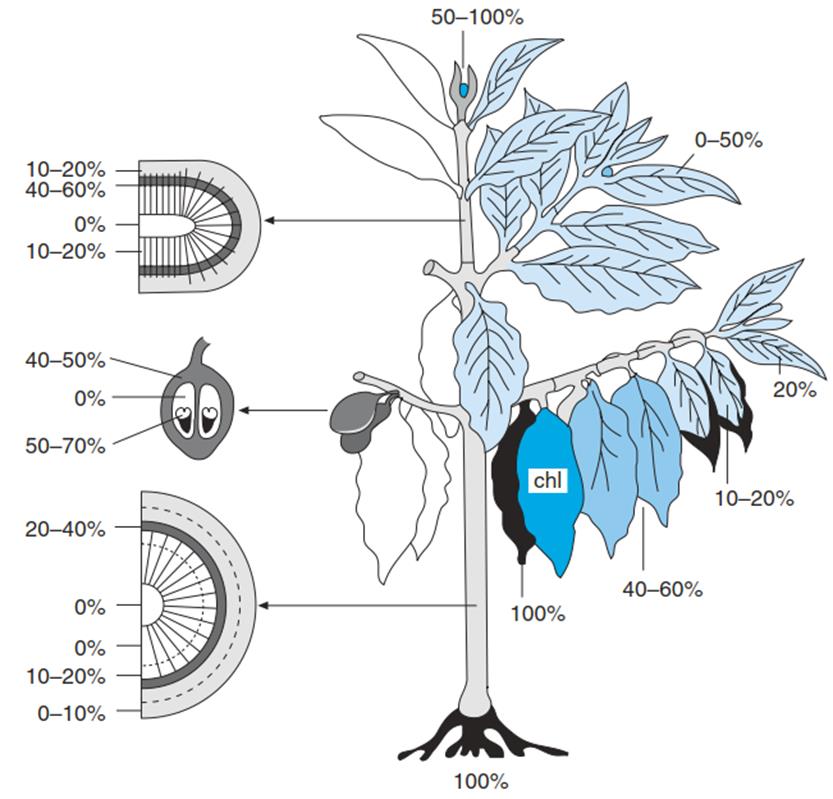
Fig. 4.9. Differences in the cold sensitivity of the organs of a coffee tree. Visual damage after 3 days of continuous cooling to +1 °C is shown. Black denotes complete damage. Chl chlorotic (Larcher 2003)
The chilling sensitivity of the African violet has been used to demonstrate that the strength or quantity of stress is composed of its intensity and its duration (Fig. 4.8). Chilling damage, visible as necroses, starts at temperatures below 8 °C. Fifty per cent damage results from 6 h of exposure to +1 °C or maintenance at +5 °C for 50 h. In this case, the low rate of repair allows comparative quantification of stress damage.
The variation in the ability to tolerate temperature extremes is far less pronounced at the other end of the temperature scale. Critical temperatures causing heat damage barely vary between plants, and hardly any correlation with the climatic zone of origin is apparent (Tables 4.3 and 4.4).
Not all life processes of a plant have the same optimum temperature, because the individual developmental steps may take place in different ambient conditions. Winter cultivars of cereals, for instance, germinate at temperatures considerably lower than those required for growth (in spring and summer). Nevertheless, the rates of germination show typical temperature dependence with Q10 values between 1.5 and 2.5. In that context it is also important to mention that not all organs of a higher plant are similarly cold sensitive or tolerant. Those most sensitive are the roots and meristems of the shoots, while axial tissues and mature leaves are the most tolerant parts (Fig. 4.9). Also, certain stages in the development of a plant are particularly vulnerable—namely, seed germination and fruit ripening.
Also relevant for plant survival is the fact that the temperature tolerance of a cell is strongly influenced by its water status. When dry, poikilohydric plants, prokaryotes and fungi can survive freezing in liquid nitrogen and exposure to temperatures close to 100 °C, while in the hydrated state the range of their temperature tolerance is much narrower. Table 4.3 presents some examples of the temperature tolerance of poikilohydric lower and higher plants, bacteria and fungi in the dry state and the hydrated state.
The correlation between the cold/frost tolerance of a species and the average minimum temperature of the climatic zone it inhabits shows the adaptive importance of mechanisms to cope with temperatures near and especially below the freezing point. Likewise, within-species variation in frost tolerance is associated with distribution of ecotypes along temperature gradients. The frost tolerance of Arabidopsis thaliana accessions correlates well with the mean minimum temperature at the site of origin. Individuals from Sweden or Russia, for instance, survive exposure to two consecutive nights of -10 °C much better than individuals from Spain, Libya or Morocco (Zhen and Ungerer 2008, Fig. 4.10). Similarly, there are pronounced differences in freezing tolerance between winter cereals and spring cereals.
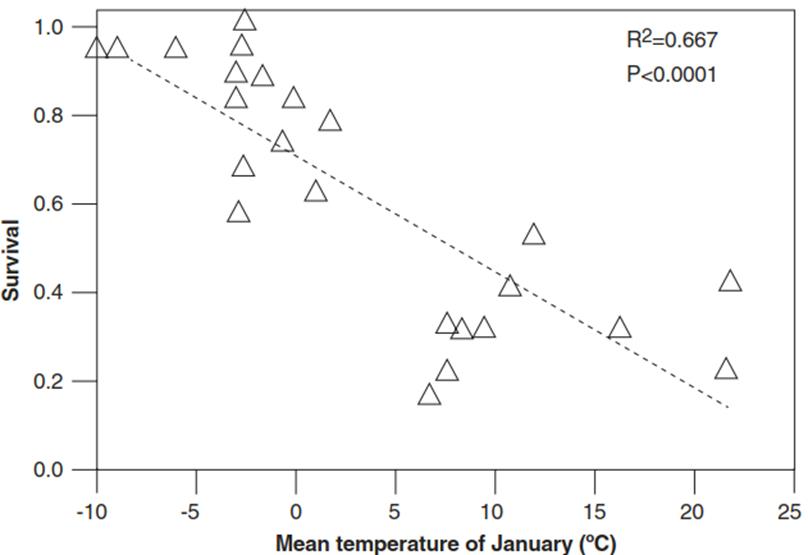
Fig. 4.10. Clinal variations in the freezing tolerance of Arabidopsis thaliana accessions. Survivorship is plotted against the January mean temperature at the site of origin for 24 A. thaliana accessions cooled to -10 °C for two consecutive nights. All plants were first cold acclimated for 7 days at 4 °C. The data are based on 20 replicates per accession (Zhen and Ungerer 2008)
Depending on the temperature climate of their habitat, plants must be adapted to and acclimate to not only the extremes of the temperature range (i.e. to seasonal variations) but also to smaller or greater diurnal temperature fluctuations. Considerable diurnal temperature oscillations are typical of deserts and the alpine and nival vegetation belts of high mountains, especially in the tropics. The air temperature during daytime may reach 20-30 °C, whereas the nocturnal temperature usually falls below the freezing point and the leaves may be stiffly frozen at the end of the night. While freezing of leaf tissue water is a process of several hours, thawing takes place within minutes once the tropical morning sun hits the leaves. These plants have developed mechanisms to slow down freezing or even avoid it. They have to be frost tolerant all year round.
On a sunny day in temperate regions (e.g. in Central Europe) the surface of bare soil may become as hot as +50 °C, while the air temperature above the soil decreases rapidly with the distance from the soil surface. The gradient in the soil itself is even steeper. Reverse temperature gradients occur also during clear nights when no cloud cover mitigates radiation emission from horizontal and slightly inclined surfaces of the vegetation or from the soil. Young plants whose shoots or leaves are close to the ground are therefore exposed to a stronger temperature stress during both day and night than plants whose leaves are supported by a stem, which experience a considerably less challenging temperature microclimate.
Date added: 2025-01-17; views: 459;