Acquired Thermotolerance
It is well documented that exposure to either a continuously applied non-lethal heat stress or single heat shocks results in the acquisition of thermotolerance or, in other words, heat hardening. These types of heat exposure elicit cellular processes that enhance the ability of plants to survive a later and more severe heat stress. Acquired thermotolerance therefore represents a classic acclimation. After just 15 min of exposure to heat, plants start to acclimatise, as indicated by a steady rise in the maximum tolerated temperatures (Kaplan et al. 2004).
Fig. 4.28 shows the differences in survival rates of A. thaliana seedlings after heat stress of 45 °C, depending on the temperature exposure prior to the heat stress.
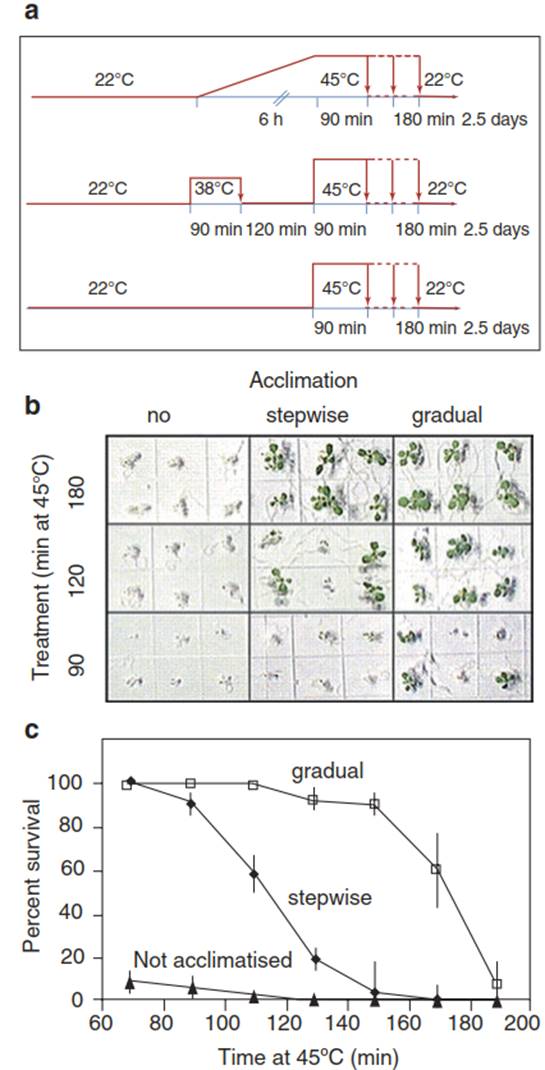
Fig. 4.28. Effect of hardening on the survival of heat-stressed Arabidopsis thaliana seedlings. a Protocol of the hardening experiment. Gradual hardening was accomplished by slow elevation of the ambient temperature from 22 °C to the heat shock temperature of 45 °C. Stepwise hardening was performed by application of a moderate heat shock (38 °C for 90 min) 2 h prior to the actual heat shock. No hardening treatment was applied in the control set of seedlings. Samples were returned to the ambient temperature and analysed for survival after 2½ days b. (c) Quantitative analysis of the acclimation effect. (Modified from Larkindale and Vierling (2008))
Similarly, heat exposure is less damaging to the photosynthetic capacity of fir needles when they have been exposed to a previous non-lethal heat stress (Fig. 4.29).
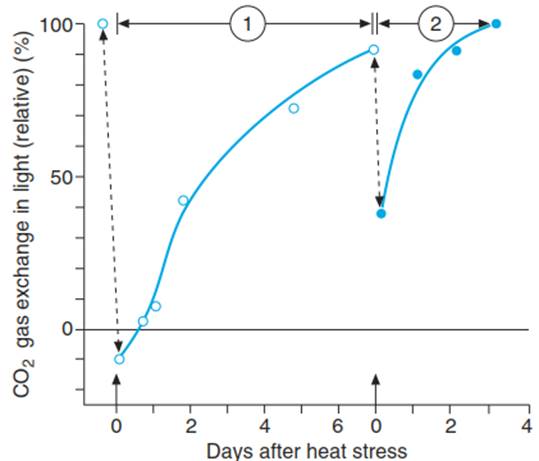
Fig. 4.29. Acquisition of thermotolerance in fir needles after heat stress treatment. Damage and hardening were assessed by measurement of the photosynthetic activity of the needles. (1) Recovery and hardening phase after the first heat stress. (2) After the second heat treatment (44 °C for 30 min). The extent of damage and the duration of the recovery phase are clearly reduced after the second stress (Larcher (1987))
The most important mechanism underlying acquired thermotolerance is the massive production of heat shock proteins (HSPs) (Sect. 4.3.3.1) (Vierling 1991). Regardless of the optimal growing temperature, this response is highly conserved not only among plants but also among prokaryotes, animals and fungi. According to recent genome-wide analyses in A. thaliana, however, not only the expression level of HSP genes changes.
Up to 2% of all genes are affected in their activity (Kotak et al. 2007). For many of them the contribution to thermotolerance is functionally not understood. What is evident is an up-regulation of anti-oxidative defences to counteract the increased production of ROS under heat stress. Documented is also the accumulation of so-called chemical chaperones, which can increase the heat resistance of cells and shift the onset of the heat stress response to higher temperatures.
These chemical chaperones are compatible solutes such as glycerol, proline and betaines. They stabilise the folding and structure of mature proteins, and they can buffer the cellular redox potential. Another critical response is the modulation of membrane lipids towards a higher saturation level of fatty acids. Some of the reactions may initiate signalling cascades that result in the induction of HSPs. For instance, the dehydration-responsive transcription factor DREB2A regulates the heat shock factor Hsf3A. ABA concentrations rise in heat-stressed tissues.
Date added: 2025-01-18; views: 581;